Icteroidae Brehm, 1828
If you're paying attention to the names of the higher-level groups, this one may leave you scratching your head. What's Icteroidae? Is it a typo for Icteroidea?
It is not a typo, but is the ending for an epifamily, a rarely used group between family and superfamily. I've resorted to it here to emphasize that the following families are more closely related to each other than to anything else.
Sibley and Monroe (1990) used the name “Emberizinae” for the remaining species. They were using an absolute measure of genetic distance to separate families and other groups. By the time you get here, everything is closely enough related to fit in a single Sibley-Monroe subfamily. In fact, even all of the nine-primaried oscines end up in the same family!
I use terms such as order and family in an ordinal, not cardinal, fashion. This lets me refer to the Sibley-Monroe “Emberizinae” as an epifamily. However, the correct name is Icteroidae (Vigors, 1825), not Emberizoidae (Brehm, 1828). This gives me a little more headroom, but as you will see below, I'm still running out of levels.
All but two of the families in Icteroidae are restricted to New World. The exceptions are Calcariidae and Emberizidae. The latter may have its origin in the New World, but is restricted to the Old World.
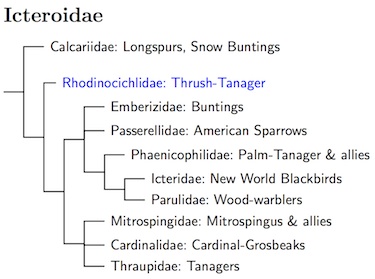
The topology of the remaining Icteroidae families is still unclear. Klicka et al. (2003) had Parulidae sister to Passerellidae, while Barker et al. (2013) give a species tree showing Emberizidae and Passerellidae as sisters, but their concatenated tree shows Emberizidae basal and Passerellidae sister to the blackbird/warbler clade. Klicka et al. (2007) has Emberizidae and Passerellidae sister, with Icteridae sister to that, and Parulidae sister to the rest.
The fact that Barker et al.'s (2013) study was unable to find definitive answers as to how these families are related was not due to lack of trying. They analyzed DNA from 6 genes and every genus in Icteroidae (genera as given by the Howard-Moore checklist (Dickinson, 2003)). Although the standard families were well-supported, this cannot be said about their relationships or inner structure. Moreover, a number of taxa did not consistently fall into any of the standard Icteroidae families: Rhodinocichla, Icteria, Teretistris, Zeledonia, Phaenicophilidae, and Mitrospingidae.
The basic problem seems to be very rapid diversification. All of Icteroidae, about 8% of all bird species, share a recent common ancestor. Barker et al. estimate this ancestor lived roughly 15 million years ago and some of the families date from a mere 10 million years ago. With such recent diversification, issues such as speciation before lineages can sort out make it particularly difficult to reconstruct the actual species tree from inconsistent gene trees. More data will be needed to completely sort this out, and it will probably not happen quickly.
 |
| TiF Icteroidae Phylogeny |
|---|
The TiF list now uses a compromise phylogeny based on the two Barker et al. (2013) estimates using all data. The Calcariidae are basal. The relationship between the Emberizidae and Passerellidae is left unresolved. The remaining Icteroidae are arranged in two groups, a clade containing the blackbirds and warblers and some Antillean Tanagers, and a thraupid clade consisting of the cardinals, tanagers, and Mitrospingidae. The exact limits of most of these families have been the subject of a lot of research. Although we mostly know the boundaries of these families, we are still uncertain about where a few genera go. Besides the aforementioned Rhodinocichla, the genera Teretistris and Zeledonia are not currently assigned to families, although Teretistris and Zeledonia belong to the blackbird/warbler clade.
Calcariidae: Longspurs, Snow Buntings Ridgway, 1901
2 genera, 6 species Not HBW Family
 |
| Calcariidae tree |
|---|
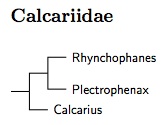
Genetic evidence shows the longspurs and snow buntings are not part of Emberizidae (buntings) or Passerellidae (American sparrows). Rather, they should be placed in their own family, Calcariidae, which is a basal branch in the Icteroidae (Klicka et al., 2003). It was also necessary to revive Rhynchophanes for McCown's Longspur in order to avoid lumping the whole family in one genus. Putting snow buntings and longspurs in one genus just didn't seem right.
The structure as found by Klicka et al. (2003) is that Rhynchophanes and Plectrophenax are sister genera, and the two together are sister to Calcarius. Maley and Wink (2010) found that McKay's Bunting, Plectrophenax hyperboreus, has only recently separated from Snow Bunting, Plectrophenax nivalis, apparently during the last glaciation. There's evidence that its population was once much larger, possibly being widespread in Beringia.
- McCown's Longspur, Rhynchophanes mccownii
- Snow Bunting, Plectrophenax nivalis
- McKay's Bunting, Plectrophenax hyperboreus
- Lapland Longspur, Calcarius lapponicus
- Smith's Longspur, Calcarius pictus
- Chestnut-collared Longspur, Calcarius ornatus
Rhodinocichlidae: Rosy Thrush-Tanager
1 genus, 1 species
The classification of the Thrush-Tanager has long been an issue. Originally considered an ovenbird by Lesson (Furnarius roseus), Hartlaub placed it in its own genus, Rhodinicichla in 1853. Since then, its classification has varied. It has been considered a type of warbler, wren, or thrasher, with names such as thrush-warbler or wren-warbler. It was eventually recognized as a nine-primaried oscine of some sort. Clark (1913) argued that it was a tanager due to similiarities with Mitrospingus, but we don't consider Mitrospingus to be a tanager. Skutch (1962) suggested it might need its own family. Eisenmann (1962) replied with an analysis meant to convince us that Rhodinocichla is a tanager. In retrospect, Eisenmann's reply is unconvincing due to the fact that many of possibly related birds he mentions are no longer part of Tharupidae.
Barker et al. (2013) is the only paper that analyzes DNA from the Rosy Thrush-Tanager. Their results are rather inconclusive. The species tree places it sister to Calcariidae, although this is rather hard to swallow. The concantenated gene tree puts it sister to the remaining Icteroidae. The mitochondrial tree puts it sister to Thraupidae. Support for any of these options is weak and I was intending to leave it unclassified even though Barker et al. suggest treating it as a separate family, Rhodinocichlidae. The discussions in Clark (1913), Eisenmann (1962), and Skutch (1962) convinced me this really is a unique taxon, and that it deserves family status.
The thrush-tanager's range is somewhat disjoint. Ridgway (1902) considered the Mexican race a distinct species. The Central and South American forms are distinct, and may also be separate species.
- Rosy Thrush-Tanager, Rhodinocichla rosea
Buntings and Sparrows
Emberizidae: Buntings Brehm, 1828
4 genera, 44 species HBW-16
The name Emberizidae is often attributed to Vigors (1825), but he did not establish the term. The attribution seems to be based on his use of a plural form of Emberiza (Emberizae), which was not used as a family-group name (Olson, 1995). It is sometimes cited as Brehm, 1831. However, Laurent Raty found an earlier use by Brehm (1828 in Isis von Oken vol. 28, Heft XII, p. 1278). Although Brehm attributes it to Vigors, there is no citation. In 1831, in his “Handbuch der Naturgeschichte aller Vogel Deutschlands”, Brehm attributed it to Vigors in the text (p.289), but to Boie in the index (p.1049). I'm guessing these all refer to the use of Emberizae mentioned above, so I attribute Emberizidae to Brehm.
 |
| Click for species-level tree for Emberizidae |
|---|
There is some uncertainty concerning whether the Emberiza buntings and American sparrows are sister clades, and that is why I have them in separate families. For example, Klicka et al. (2003) has Emberizidae embedded in the Icteridae while Passerellidae is sister to Parulidae.
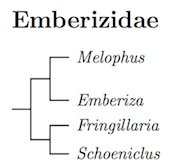
Taxonomy within Emberizidae now follows Päckert et al. (2015). The major clades are similar to those found by Alström et al. (2008a) and the adjustments based on Ren et al. (2014) and Schweizer and Kirwan (2014). However, the arrangement of the clades is a bit different, and is not as well supported as I would like.
As in H&M-4 (Dickinson and Christidis, 2014), I have treated the major clades as separate genera. The only exception is that I do not recognize Granativora. With the Päckert et al. topology it would require a separate genus for the Brown-rumped Bunting, Melophus affinis, and no such name is available. The position of the Brown-rumped Bunting is based on limited genetic data and support is low, so it may belong elsewhere.
Should anyone feel inclined to further divide these genera, there are available names for some of the subclades of these clades. For one such possibility, see the species-level tree for Emberiza. The groupings are in general agreement with the morphological groups of Byers et al. (1995). Their Yellowhammer group is subgenus Emberiza; the Rock Bunting group is Cia; the Ortolan Bunting group is Glycyspina; the House Bunting and Golden-breasted bunting groups are both clades within Fringillaria. Not all of Byers et al.'s species pairs are supported.
Sharpe's Bunting, Emberiza yunnanensis (probably including khamensis), has been split from Godlewski's Bunting, Emberiza godlewskii (see Päckert et al., 2015). There didn't seem to be historical English name to press into service. I chose Sharpe as he named yunnanensis. Yunnan Bunting seemed a little limiting because the range extends into Sichuan, and via khamensis onto the Tibetian plateau.
Gosling's Bunting, Fringillaria goslingi, has been split from Cinnamon-breasted Bunting, Fringillaria tahapisi. See Olsson et al. (2013b).
- Brown-rumped Bunting, Melophus affinis
- Crested Bunting, Melophus lathami
- Black-headed Bunting, Melophus melanocephalus
- Red-headed Bunting, Melophus bruniceps
- Corn Bunting, Emberiza calandra
- Chestnut-eared Bunting, Emberiza fucata
- Tibetan Bunting, Emberiza koslowi
- Jankowski's Bunting, Emberiza jankowskii
- Rock Bunting, Emberiza cia
- Sharpe's Bunting, Emberiza yunnanensis
- Meadow Bunting, Emberiza cioides
- Godlewski's Bunting, Emberiza godlewskii
- Gray-necked Bunting, Emberiza buchanani
- Cinereous Bunting, Emberiza cineracea
- Ortolan Bunting, Emberiza hortulana
- Cretzschmar's Bunting, Emberiza caesia
- Cirl Bunting, Emberiza cirlus
- White-capped Bunting, Emberiza stewarti
- Yellowhammer, Emberiza citrinella
- Pine Bunting, Emberiza leucocephalos
- Cabanis's Bunting, Fringillaria cabanisi
- Golden-breasted Bunting, Fringillaria flaviventris
- Somali Bunting, Fringillaria poliopleura
- Cape Bunting, Fringillaria capensis
- Lark-like Bunting, Fringillaria impetuani
- Socotra Bunting, Fringillaria socotrana
- House Bunting, Fringillaria sahari
- Striolated Bunting, Fringillaria striolata
- Gosling's Bunting, Fringillaria goslingi
- Cinnamon-breasted Bunting, Fringillaria tahapisi
- Yellow-throated Bunting, Schoeniclus elegans
- Slaty Bunting, Schoeniclus siemsseni
- Ochre-rumped Bunting / Japanese Reed-Bunting, Schoeniclus yessoensis
- Reed Bunting / Common Reed-Bunting, Schoeniclus schoeniclus
- Pallas's Bunting / Pallas's Reed-Bunting, Schoeniclus pallasi
- Yellow-breasted Bunting, Schoeniclus aureolus
- Rustic Bunting, Schoeniclus rusticus
- Little Bunting, Schoeniclus pusillus
- Black-faced Bunting, Schoeniclus spodocephala
- Yellow Bunting, Schoeniclus sulphuratus
- Chestnut Bunting, Schoeniclus rutilus
- Yellow-browed Bunting, Schoeniclus chrysophrys
- Tristram's Bunting, Schoeniclus tristrami
- Gray Bunting, Schoeniclus variabilis
Passerellidae: American Sparrows Cabanis & Heine, 1850-51
28 genera, 138 species HBW-16 (split)
 |
| Click for genus-level tree for Passerellidae |
|---|
The sparrow family has been carved up in the ongoing reorganization of the nine-primaried oscines. Most of the Neotropical finches have joined the tanagers. The Gubernatrix and Paroaria cardinals have been moved to the tanagers. The Neotropical finches that remain are the Atlapetes brushfinches, as well as the Large-footed, Yellow-thighed, and Yellow-green Finches (Pezopetes and Pselliophorus. In return, the sparrows gain the genus Chlorospingus (excepting flavovirens) and the Tanager Finch, Oreothraupis arremonops. These changes alone would reduce the family to about half its former size, but there is more. They also lose the Emberiza.
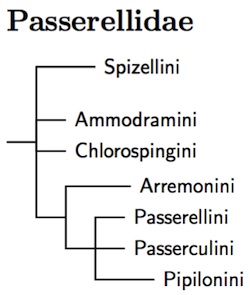
What remains is a somewhat more homogeneous family and it is possible to give it a coherent organization. Everything seems to fall into 8 clades, most of which I've treated at the tribe level (I have united two of them). I haven't used subfamilies partly because I don't think it's clear yet how the tribes fit together, and partly because I don't think the divisions are particularly deep.
The correct arrangement of the Passerellidae tribes remains unclear. The version presented here is based on Klicka et al. (2014) and is similar to previous arrangements based on Barker et al. (2013), which has Spizellini as the basal group. There is enough uncertainty (Carson and Spicer, 2003, DaCosta et al., 2009, and Klicka et al., 2007 all have slightly different arrangements) that I leave it as part of a basal trichotomy. All put the remaining tribes in the third branch of the trichotomy.
We consider the Spizellini first. As shown by Carson and Spicer (2003), this clade includes most of the Spizella sparrows. The genus Amphispiza is restricted to the Five-striped Sparrow and the Black-throated Sparrow, but does not include the sage sparrows. The arrangement within Spizella follows Canales-Del Castillo et al. (2010), with Brewer's Sparrow somewhat surprisingly sister to Worthen's Sparrow. The Timberline Sparrow, Spizella breweri taverneri, is sometimes thought to be a separate species. Although there is some differentiation between it and Brewer's Sparrow, it may be best thought of as an incipient species within Brewer's Sparrow. See Klicka et al. (1999).
This brings us to Ammodramini, where some genera have required reorganization. Ammodramus and Aimophila have been affected the most. The papers by Klicka and Spellman (2007) and DaCosta et al. (2009) show what to do. Ammodramus itself is reduced to the Grasshopper Sparrow together with a couple of Neotropical relatives. These are sister to Arremonops. Together, they are sister to a big chunk of what was Aimophila. As the Aimophila type species is not included, I have revived Audubon's 1839 name for them, Peucaea (type aestivalis). Two more of the former Aimophila are sister to the rest of Ammodramini. Following DaCosta et al.'s suggestion, they take the genus name Rhyncospiza (Ridgway 1898, type stolzmanni).
The other big group in Ammodramini consists of a some Neotropical sparrows. These have been all merged into the genus Arremon. Although there are 4 distinct groups, the actual branching order remains a bit unclear (see Cadena et al., 2007 and Flórez-Rodríguez et al., 2011). One of the leading candidates is illustrated on the tree.
The groups are the two former Lysurus finches, the brunneinucha/virenticeps complex, which may actually hide a half-dozen species (Navarro-Sigüenza et al., 2007), the torquatus group, which has been split into 8 species (Cadena et al., 2007, 2010, 2011; also see SACC proposal #468), and the Arremon sparrows. The split of the Stripe-headed Brushfinch (torquatus group) is detailed below. The English names follows AOU's NACC and SACC
| The Stripe-headed Brushfinch Complex | ||
|---|---|---|
| Species | Subspecies | Location |
| Costa Rican Brushfinch | costaricensis | Costa Rica, W Panama |
| Gray-browed Brushfinch | assimilis, larensis, nigrifrons, poliophrys | Andes of Venezuela, Colombia, Ecuador, Peru |
| Sierra Nevada Brushfinch | basilicus | Santa Marta Mountains |
| Perija Brushfinch | perijanus | E Colombia to W Venezuela |
| White-browed Brushfinch | torquatus, fimbriatus, borelli | extreme S Peru, Bolivia, Argentina |
| Black-headed Brushfinch | atricapillus, tacarcunae | C & E Panama, Colombian Andes |
| Caracas Brushfinch | phaeopleurus | Coastal mountains of N Venezuela |
| Paria Brushfinch | phygas | NE Venezuela |
Chlorospingini consists of Chlorospinus and the Tanager Finch (Oreothraupis) Although it had been suggested that the Tanager Finch belongs with the Atlapetes Brushfinches, Barker et al. (2013) found it sister to Chlorospingus. The treatment of the Common Chlorospingus complex is based on García-Moreno et al. (2004), Sánchez-González et al. (2007), Bonaccorso et al. (2008), and Weir et al. (2008). This involves breaking up the Common Chlorospingus into 9 species. Four of these are primarily Mexican: White-fronted Chlorospingus, C. albifrons; Wetmore's Chlorospingus, C. wetmorei; Brown-headed Chlorospingus, C. ophthalmicus; and Dwight's Chlorospingus, C. dwighti. The color plate in Sánchez-González et al. (2007) illustrates these forms as well as Dusky-headed Chlorospingus, C. postocularis. Other taxa that appear to deserve species status are the Central American Dotted Chlorospingus, C. punctulatus; the Buff-breasted Chlorospingus, C. cinereocephalus, of Peru; and the Yellow-breasted Chlorospingus, C. flavopectus. The list of subspecies below presumes that Isler and Isler's (1987) flavopectus group stays together, although only two of its subspecies were analyzed by Weir et al. (2008). I've also presumed that Isler and Isler's venezuelanus group stays together. Weir found that some of them group with several more southern races. For the present, it seems reasonable to put the whole lot of them in a single species and call it Common Chlorospingus, C. venezuelanus. There may still be additional species hiding within the Common Chlorospingus complex.
| The Common Chlorospingus Complex | ||
|---|---|---|
| Species | Subspecies | Location |
| Wetmore's Chlorospingus | wetmorei | Mexico: Sierra de Tuxtla |
| White-fronted Chlorospingus | albifrons, persimilis | Mexico: Sierra Madre del Sur |
| Brown-headed Chlorospingus | ophthalmicus | Mexico: Sierra Madre Oriental |
| Dwight's Chlorospingus | dwighti | Mexico: Chiapas, Caribbean slope |
| Dusky-headed Chlorospingus | postocularis, honduratius | Chiapas, Pacific slope; Guatemala, Honduras, El Salvador |
| Dotted Chlorospingus | punctulatus, regionalias, ‘novicius’* | Nicaragua, Costa Rica, W. Panama |
| Common Chlorospingus | venezuelanus*, jacqueti, falconensis*, ponsi*, eminens*, peruvianus, bolivianus, fulvigularis, argentinus | E. Colombia, Venezuela, S. Peru, Bolivia, Argentina |
| Buff-breasted Chlorospingus | cinereocephalus | C. Peru |
| Tacarcuna Chlorospingus | tacarcunae | E. Panama |
| Pirre Chlorospingus | inornatus | Darién (Panama, Colombia) |
| Yellow-breasted Chlorospingus | flavopectus*, trudis*, exitelis*, macarenae*, nigriceps*, phaeocephalus, hiaticolus | Colombia, Ecuador, N. Peru |
| Dusky Chlorospingus | semifuscus, livingstoni* | W. Colombia, W. Ecuador |
| Subspecies based on Dickinson et al. (2003). *An asterisk indicates taxa not sampled by Weir et al. (2008). The subspecies ‘novicius’ may be a hybrid form. | ||
The third group of sparrows includes three tribes: Passerellini, Passerculini, and Pipilonini. Although there is consensus that they are each other's closest relatives, it remains unclear exactly how they relate. Barker et al. (2013) supports treating Passerellini and Passerculini as sisters, as is done here.
The Junco/Zonotrichia clade Passerellini is first. It is now clear that the American Tree Sparrow is not part of Spizella. Rather, it is most closely related to the Fox Sparrow, Passerella iliaca (Carson and Spicer, 2003). Slager and Klicka (2014a) established the new genus Spizelloides for it. The Passerella/Spizelloides group forms a clade with the juncos and Zonotrichia. Passerella iliaca itself remains controversial, and may end up being split into four species. The Guadalupe Junco, Junco insularis, has been split from J. hyemalis (it is actually sister to hyemalis + phaeonotus, see Klicka et al., 2014).
The rest of Ammodramus ends up in the Passerculini. The “marshland” Ammodramus form a clade. As Klicka and Spellman (2007) recommend, they get the name Ammospiza. Although Henslow's and Baird's Sparrows may be each other's closest relatives, it is also possible they are not. Since there is only one available genus name between them, I've put them both in Centronyx (Baird 1858, type bairdii). Note that Nemospiza is preoccupied by an arachnid, and is not available. The sage sparrows get a brand new genus name. Klicka and Spellman (2007) discovered that it is not related to the other Amphispiza. The name they proposed, Artemisospiza, did not strictly follow ICZN rules. In view of this, Klicka and Banks (2011) proposed the name Artemisiospiza (think sagebrush, Artemesia). Moreover, the AOU, after considering Cicero and Koo (2012), now treats the Sage Sparrow as two species: Bell's Sparrow, Artemisiospiza belli, and Sagebrush Sparrow, Artemisiospiza nevadensis. The situation is rather confusing and a third species may be involved.
DaCosta et al. (2009) resolved a big chunk of the uncertainty concerning the Pipilonini. The Melozone Ground-Sparrows end up in two (or three?) separate groups. One part is sister to the brown towhees. DaCosta et al. suggest the genus name Pyrgisoma for it. The AOU proposal suggested the type is kieneri, although Ridgway gives the type as biarcuata. Apparently, treating kieneri as type dates from a time when biarcuata was considered a subspecies of kieneri. However, when separate, kieneri becomes the type species of Kieneria (Bonaparte 1855) and biarcuata is the type of Pyrgisoma. Since biarcuata ends up in the genus Melozone, Kieneria is the name used here. The brown towhees have always been considered different, and there have been suggestions they should get their own genus. That has never happened, so my best option seems to be to put them into Kieneria too. Surprisingly, the Zapata Sparrow, Torreornis inexpectata, thought to be sister to Amphispiza, turns out to be sister to Kieneria (Barker et al., 2013).
The other part of Melozone is merged into Aimophila because they are apparently successive sisters to the core Aimophila. This is surprising as a look at them makes it seem they must be sisters. Barker et al. (2013) found the Large-footed Finch, Pezopetes capitalis, to be basal to both, although support for this was not great.
The rest of the Pipilonini consists of the Pipilo towhees together with the Atalapetes brushfinches and allies. The extinct Bermuda Towhee, Pipilo naufragus, has been included as it seems to have survived into historical times. See Olson and Wingate (2012).
The Pselliophorus are embedded in Atlapetes, so I have changed their scientific names according. Klicka et al. (2014) present a comprehensive molecular phylogeny of Atlapetes which includes almost all of the Atlapetes species. The linear order is based on their phylogeny. One species they left out was the Black-faced Brushfinch, Atlapetes melanolaemus. Its position is based on Sánchez-González et al. (2015).
Based on Sánchez-González et al. (2015), the Choco Brushfinch, Atlapetes crassus (Colombia and Ecuador) has been split from Tricolored Brushfinch, Atlapetes tricolor (Peru), and Taczanowski's Brushfinch, Atlapetes taczanowskii (Peru), has been split from Slaty Brushfinch, Atlapetes schistaceus (Colombia, Ecuador, and Venezuela).
Based on Donegan et al. (2014a), Merida Brushfinch, Atlapetes meridae, has been split from Moustached Brushfinch, Atlapetes albofrenatus, and Black-fronted Brushfinch, Atlapetes nigrifrons, has been split from Yellow-breasted Brushfinch, Atlapetes latinuchus. A further split from A. latinuchus may be needed, but it is not yet clear how that will work.
Spizellini Baird, 1858
- Lark Sparrow, Chondestes grammacus
Click for species-level
Passerellidae tree - Lark Bunting, Calamospiza melanocorys
- Black-throated Sparrow, Amphispiza bilineata
- Five-striped Sparrow, Amphispiza quinquestriata
- Chipping Sparrow, Spizella passerina
- Clay-colored Sparrow, Spizella pallida
- Black-chinned Sparrow, Spizella atrogularis
- Field Sparrow, Spizella pusilla
- Brewer's Sparrow, Spizella breweri
- Worthen's Sparrow, Spizella wortheni
Ammodramini: Grassland Sparrows Ridgway, 1901
- Tumbes Sparrow, Rhynchospiza stolzmanni
- Stripe-capped Sparrow, Rhynchospiza strigiceps
- Grasshopper Sparrow, Ammodramus savannarum
- Grassland Sparrow, Ammodramus humeralis
- Yellow-browed Sparrow, Ammodramus aurifrons
- Olive Sparrow, Arremonops rufivirgatus
- Green-backed Sparrow, Arremonops chloronotus
- Black-striped Sparrow, Arremonops conirostris
- Tocuyo Sparrow, Arremonops tocuyensis
- Rufous-winged Sparrow, Peucaea carpalis
- Cinnamon-tailed Sparrow, Peucaea sumichrasti
- Botteri's Sparrow, Peucaea botterii
- Cassin's Sparrow, Peucaea cassinii
- Bachman's Sparrow, Peucaea aestivalis
- Stripe-headed Sparrow, Peucaea ruficauda
- Black-chested Sparrow, Peucaea humeralis
- Bridled Sparrow, Peucaea mystacalis
Chlorospingini Informal
- Tanager Finch, Oreothraupis arremonops
- Short-billed Chlorospingus / Yellow-whiskered Bush-Tanager, Chlorospingus parvirostris
- Yellow-throated Chlorospingus, Chlorospingus flavigularis
- Ashy-throated Chlorospingus, Chlorospingus canigularis
- Sooty-capped Chlorospingus, Chlorospingus pileatus
- Wetmore's Chlorospingus, Chlorospingus wetmorei
- White-fronted Chlorospingus, Chlorospingus albifrons
- Brown-headed Chlorospingus, Chlorospingus ophthalmicus
- Dwight's Chlorospingus, Chlorospingus dwighti
- Dusky-headed Chlorospingus, Chlorospingus postocularis
- Dotted Chlorospingus, Chlorospingus punctulatus
- Common Chlorospingus, Chlorospingus venezuelanus
- Buff-breasted Chlorospingus, Chlorospingus cinereocephalus
- Tacarcuna Chlorospingus, Chlorospingus tacarcunae
- Pirre Chlorospingus, Chlorospingus inornatus
- Yellow-breasted Chlorospingus, Chlorospingus flavopectus
- Dusky Chlorospingus, Chlorospingus semifuscus
Arremonini: Scrub Sparrows Sundevall, 1872
- Sooty-faced Finch, Arremon crassirostris
- Olive Finch, Arremon castaneiceps
- Green-striped Brushfinch, Arremon virenticeps
- Chestnut-capped Brushfinch, Arremon brunneinucha
- Orange-billed Sparrow, Arremon aurantiirostris
- Black-capped Sparrow, Arremon abeillei
- Golden-winged Sparrow, Arremon schlegeli
- Saffron-billed Sparrow, Arremon flavirostris
- Pectoral Sparrow, Arremon taciturnus
- Sao Francisco Sparrow, Arremon franciscanus
- Half-collared Sparrow, Arremon semitorquatus
- Costa Rican Brushfinch, Arremon costaricensis
- Paria Brushfinch, Arremon phygas
- Black-headed Brushfinch, Arremon atricapillus
- Caracas Brushfinch, Arremon phaeopleurus
- Gray-browed Brushfinch, Arremon assimilis
- White-browed Brushfinch, Arremon torquatus
- Sierra Nevada Brushfinch, Arremon basilicus
- Perija Brushfinch, Arremon perijanus
Passerellini: Juncos and allies Cabanis & Heine, 1850-51
- American Tree Sparrow, Spizelloides arborea
- Fox Sparrow, Passerella iliaca
- Volcano Junco, Junco vulcani
- Guadalupe Junco, Junco insularis
- Dark-eyed Junco, Junco hyemalis
- Yellow-eyed Junco, Junco phaeonotus
- Rufous-collared Sparrow, Zonotrichia capensis
- Harris's Sparrow, Zonotrichia querula
- White-throated Sparrow, Zonotrichia albicollis
- Golden-crowned Sparrow, Zonotrichia atricapilla
- White-crowned Sparrow, Zonotrichia leucophrys
Passerculini Informal
- Bell's Sparrow, Artemisiospiza belli
- Sagebrush Sparrow, Artemisiospiza nevadensis
- Striped Sparrow, Oriturus superciliosus
- Vesper Sparrow, Pooecetes gramineus
- Le Conte's Sparrow, Ammospiza leconteii
- Seaside Sparrow, Ammospiza maritima
- Nelson's Sparrow, Ammospiza nelsoni
- Saltmarsh Sparrow, Ammospiza caudacuta
- Savannah Sparrow, Passerculus sandwichensis
- Baird's Sparrow, Centronyx bairdii
- Henslow's Sparrow, Centronyx henslowii
- Sierra Madre Sparrow, Xenospiza baileyi
- Lincoln's Sparrow, Melospiza lincolnii
- Song Sparrow, Melospiza melodia
- Swamp Sparrow, Melospiza georgiana
Pipilonini: Towhees & BrushfinchesBonaparte, 1854
- Large-footed Finch, Pezopetes capitalis
- Zapata Sparrow, Torreornis inexpectata
- Rusty-crowned Ground-Sparrow, Kieneria kieneri
- Canyon Towhee, Kieneria fusca
- White-throated Towhee, Kieneria albicollis
- California Towhee, Kieneria crissalis
- Abert's Towhee, Kieneria aberti
- White-eared Ground-Sparrow, Aimophila leucotis
- Prevost's Ground-Sparrow, Aimophila biarcuata
- Rusty Sparrow, Aimophila rufescens
- Rufous-crowned Sparrow, Aimophila ruficeps
- Oaxaca Sparrow, Aimophila notosticta
- Collared Towhee, Pipilo ocai
- Green-tailed Towhee, Pipilo chlorurus
- Spotted Towhee, Pipilo maculatus
- Eastern Towhee, Pipilo erythrophthalmus
- Bermuda Towhee, Pipilo naufragus
- Rufous-capped Brushfinch, Atlapetes pileatus
- White-naped Brushfinch, Atlapetes albinucha
- Yellow-thighed Finch, Atlapetes tibialis
- Yellow-green Finch, Atlapetes luteoviridis
- Tepui Brushfinch, Atlapetes personatus
- Black-spectacled Brushfinch, Atlapetes melanopsis
- Choco Brushfinch, Atlapetes crassus
- Apurimac Brushfinch, Atlapetes forbesi
- Black-faced Brushfinch / Gray-eared Brushfinch, Atlapetes melanolaemus
- Cuzco Brushfinch, Atlapetes canigenis
- Vilcabamba Brushfinch, Atlapetes terborghi
- Bolivian Brushfinch, Atlapetes rufinucha
- Fulvous-headed Brushfinch, Atlapetes fulviceps
- Yellow-striped Brushfinch, Atlapetes citrinellus
- Moustached Brushfinch, Atlapetes albofrenatus
- Merida Brushfinch, Atlapetes meridae
- Black-fronted Brushfinch, Atlapetes nigrifrons
- Santa Marta Brushfinch, Atlapetes melanocephalus
- Ochre-breasted Brushfinch, Atlapetes semirufus
- White-headed Brushfinch, Atlapetes albiceps
- White-rimmed Brushfinch, Atlapetes leucopis
- Yellow-headed Brushfinch, Atlapetes flaviceps
- Dusky-headed Brushfinch, Atlapetes fuscoolivaceus
- Rufous-eared Brushfinch, Atlapetes rufigenis
- Tricolored Brushfinch, Atlapetes tricolor
- Slaty Brushfinch, Atlapetes schistaceus
- Taczanowski's Brushfinch, Atlapetes taczanowskii
- Pale-naped Brushfinch, Atlapetes pallidinucha
- Yellow-breasted Brushfinch, Atlapetes latinuchus
- White-winged Brushfinch, Atlapetes leucopterus
- Pale-headed Brushfinch, Atlapetes pallidiceps
- Bay-crowned Brushfinch, Atlapetes seebohmi
- Rusty-bellied Brushfinch, Atlapetes nationi
- Antioquia Brushfinch, Atlapetes blancae