Passerida Nitzsch, 1820
The Passerida are the sister group of the Corvida. There are several basal groups that branch off before we get to the heart of the Passerida. The taxonomy of Passerida has been changing rapidly and most current checklists lag well behind the latest research.
Exactly which families are basal Passerida is still being clarified, with families being shuffled between the Passerida and Corvida. The latest on this is Aggerbeck et al. (2014). Their arrangement is similar to Irestedt and Ohlson (2008). Irestedt and Ohlson (2008) made a strong argument that several families thought to be Corvida are actually basal Passerida. Irestedt and Ohlson identified a genetic marker that seems to separate the Passerida from the Corvida. If this is correct, it means that taxonomies that rely primarily on the RAG-1 gene (e.g., Baker et al., 2004), are not entirely correct. Their paper is a reminder of the importance of using several independent genes to test taxonomic hypotheses. There are other portions of the avian tree where this standard has not yet been met; those areas are particularly subject to change.
Melanocharitoidea
Melanocharitidae: Berrypeckers & Longbills Sibley & Ahlquist, 1985
4 genera, 10 species HBW-13
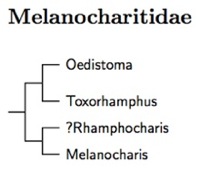 We continue with a group of New Guinea endemics. Sibley and Monroe (1993)
listed them in the Passeroidea, near the flowerpecker and sunbirds. They
include the former honeyeaters Oedistoma and Toxorhamphus.
We continue with a group of New Guinea endemics. Sibley and Monroe (1993)
listed them in the Passeroidea, near the flowerpecker and sunbirds. They
include the former honeyeaters Oedistoma and Toxorhamphus.
- Dwarf Longbill, Oedistoma iliolophus
- Pygmy Longbill, Oedistoma pygmaeum
- Yellow-bellied Longbill, Toxorhamphus novaeguineae
- Slaty-headed Longbill, Toxorhamphus poliopterus
- Spotted Berrypecker, Rhamphocharis crassirostris
- Obscure Berrypecker, Melanocharis arfakiana
- Black Berrypecker, Melanocharis nigra
- Mid-mountain Berrypecker, Melanocharis longicauda
- Fan-tailed Berrypecker, Melanocharis versteri
- Streaked Berrypecker, Melanocharis striativentris
Cnemophiloidea
The satinbirds (Cnemophilidae) have been moved here from the Corvida. These New Guinea endemics were once considered birds-of-paradise (Paradisaeidae), but now have their own family.
Cnemophilidae: Satinbirds Mayr, 1962
2 genera, 3 species Not HBW Family
- Yellow-breasted Satinbird / Silken Satinbird, Loboparadisea sericea
- Loria's Satinbird / Velvet Satinbird, Cnemophilus loriae
- Crested Satinbird / Antenna Satinbird, Cnemophilus macgregorii
Callaeoidea
The Callaeoidea include the stitchbird (Notiomystidae), the New Zealand wattlebirds (Callaeidae) and the Australasian Robins (Petroicidae). Aggerbeck et al. (2014) put Notiomystidae and Callaeidae first in Passerida. However, the complete mitchondrial analysis of Barker (2014) and the multigene (6 nuclear and 3 mitochondrial) analysis of Zuccon et al. (2012) placed them next to the Australasian Robins, as here.
The monotypic stitchbird was previously considered a honeyeater, but is actually most closely related to the wattlebirds (Ewen et al., 2006; Driskell et al., 2007). The division between the Stitchbird and other families in its clade seems quite ancient, so it is given its own family.
The genetic studies that have been done on the Australasian robins (Petroicidae) have been somewhat equivocal on its placement—whether it goes before or after the Picathartoidea. Jønsson and Fjeldså (2006a) argue that placing it first makes the best sense both genetically and biogeographically. Genetically, the rest of the Passerida share an insertion in the nuclear c-myc gene that the Petroicidae lack (Ericson et al., 2000). The biogeographic sense is that all of the Petroicidae are Australasian. It seems unlikely the remaining Passerida would develop in Africa (the Picathartoidea), then suddenly jump back to Australia before returning to Africa. Using a different set of genes, Irestedt and Ohlson (2008) also put the Australasian robins before the Picathartoidea. Recent multigene analyses such as Barker (2014) and Zuccon et al. (2012) concur.
Notiomystidae: Stitchbird Driskell et al., 2007
1 genus, 1 species HBW-14
- Stitchbird / Hihi, Notiomystis cincta
Callaeidae: New Zealand Wattlebirds Sundevall, 1836 (1831)
3 genera, 5 species HBW-14
The three wattlebird genera apparently diverged almost simultaneously (Shepherd and Lambert, 2007), and are treated as a trichotomy.
Following OSNZ, IOC, and HBW-14, the Kokako, Callaeas cinereus, is split into North Island Kokako, Callaeas wilsoni, and South Island Kokako, Callaeas cinereus. See also Double and Murphy (2002). Further, the Saddleback, Philesturnus carunculatus is split into North Island Saddleback, Philesturnus rufusater, and South Island Saddleback, Philesturnus carunculatus (see also Parker et al., 2013).
- North Island Kokako, Callaeas wilsoni
- South Island Kokako, Callaeas cinereus
- North Island Saddleback, Philesturnus rufusater
- South Island Saddleback, Philesturnus carunculatus
- Huia, Heteralocha acutirostris
Petroicidae: Australasian Robins Mathews, 1919-20
14 genera, 49 species HBW-12
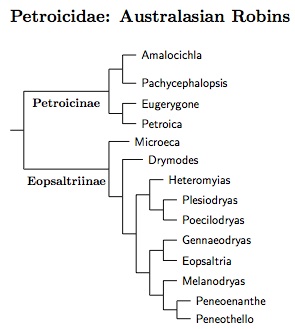 |
| Click for Petroicidae species tree |
|---|
Differences from standard arrangements of Petroicidae are based on Christidis et al. (2011), Loynes et al. (2009), and Miller and Lambert (2006). The Christidis et al. paper is particularly useful as it includes 41 of the 47 species recognized here.
Christidis et al. (2011) note six major clades in Petroicidae, and recommend that they be treated as subfamilies. I don't think this does a good job of describing the phylogeny, and prefer to separate Petroicidae into two subfamilies: Petroicinae and Eopsaltriinae. This would reduce the Christidis et al. subfamilies to tribes. In that case, the subfamily Petroicinae includes the monotypic tribes Amalocichlini (Christidis et al., 2011) and Pachycephalopsini (Christidis et al., 2011), as well as Petroicini (Eugerygone and Petroica); the subfamily Eopsaltriinae contains the monotypic Drymodini (Wolters, 1980) and Microecini (Loynes et al., 2009), with the other species grouped in Eopsaltriini.
An earlier work by Norman et al. (2009b) suggested that Amalocichla was in fact a member of Petroicidae. I had previously complained that their published analysis was inconclusive as it lacked members of Passerida. That meant that the two Amalocichla could be closer to Passer domesticus or the Stitchbird than to Petroicidae. After all, it has been previously considered part of Turdidae. Well, Christidis et al. (2011) cleared up the situation by including Passer montanus and Callaeas cinerea (close to the Stitchbird). This nails down Amalocichla as part of Petroicidae.
Based on the results of Loynes et al. (2009) and Christidis et al., the Yellow-bellied Robin has moved from Eopsaltria to Microeca. Note that Monachella ends up being submerged into Microeca. I do not follow Christidis et al.'s suggestion to divide Microeca into 5 genera.
I have restored the genus Heteromyias, which is widely recognized (but not by Dickinson et al., 2003) and placed the Black-throated Robin in Plesiodryas (Matthews, 1920), as recommended by Christidis et al.
The Banded Yellow Robin moves from Poecilodryas to the monotypic Gennaeodryas (Matthews, 1920), which is sister to Eopsaltria. Christidis et al. put the White-breasted Robin in Quoyornis in order to maintain Tregellasia. I think it makes more sense to submerge Tregellasia into Eopsaltria.
Both papers found evidence that Peneoenanthe is nested in Peneothello, although support was low. After considering the various gene trees and the species involved, I have adopted a slightly different topology that retains them in separate genera. This reverses my earlier stance on this.
Based on Miller and Lambert (2006) and H&M-4 (among others), the New Zealand Robin, Petroica australis, has been split into North Island Robin, Petroica longipes, and South Island Robin, Petroica australis (inc. rakiura). Based on Kearns et al. (2016), the Pacific Robin, Petroica multicolor, has been split into Pacific Robin, Petroica pusilla and the monotypic Norfolk Robin, Petroica multicolor.
One of the New Guinea races of Drymodes, brevirostris is no more closely related to superciliaris than to brunneopygia. I follow Christidis et al.'s suggestion to split it and the other New Guinea races as Papuan Scrub-Robin, Drymodes beccarii. They also suggest treating Melanodryas cucullata westralensis as a separate species as M. vittata otherwise renders M. cucullata paraphyletic. If this were a case of allopatric subspecies with distinct plumage or morphology, I would agree in a second. However, these are virtually indistinguishable taxa with ranges that abut one another across a good chunk of Australia. I suspect introgression here, and think that splitting them would be premature. Christidis et al. also note several other taxa in need of closer study.
Petroicinae Mathews, 1919-20
- Lesser Ground-Robin, Amalocichla incerta
- Greater Ground-Robin, Amalocichla sclateriana
- Green-backed Robin, Pachycephalopsis hattamensis
- White-eyed Robin, Pachycephalopsis poliosoma
- Garnet Robin, Eugerygone rubra
- Snow Mountains Robin, Petroica archboldi
- Mountain Robin, Petroica bivittata
- North Island Robin, Petroica longipes
- South Island Robin, Petroica australis
- Black Robin, Petroica traversi
- Tomtit, Petroica macrocephala
- Norfolk Robin, Petroica multicolor
- Red-capped Robin, Petroica goodenovii
- Pacific Robin, Petroica pusilla
- Rose Robin, Petroica rosea
- Pink Robin, Petroica rodinogaster
- Scarlet Robin, Petroica boodang
- Flame Robin, Petroica phoenicea
Eopsaltriinae Mathews, 1946
- Canary Flyrobin, Microeca papuana
- Olive Flyrobin, Microeca flavovirescens
- Yellow-legged Flycatcher / Yellow-legged Flyrobin, Microeca griseoceps
- Yellow-bellied Flyrobin, Microeca flaviventris
- Torrent Flyrobin, Microeca muelleriana
- Jacky Winter, Microeca fascinans
- Lemon-bellied Flycatcher / Lemon-bellied Flyrobin, Microeca flavigaster
- Golden-bellied Flyrobin, Microeca hemixantha
- Papuan Scrub-Robin, Drymodes beccarii
- Northern Scrub-Robin, Drymodes superciliaris
- Southern Scrub-Robin, Drymodes brunneopygia
- Ashy Robin, Heteromyias albispecularis
- Gray-headed Robin, Heteromyias cinereifrons
- Black-throated Robin, Plesiodryas albonotata
- Black-sided Robin, Poecilodryas hypoleuca
- Black-chinned Robin, Poecilodryas brachyura
- Buff-sided Robin, Poecilodryas cerviniventris
- White-browed Robin, Poecilodryas superciliosa
- Banded Yellow Robin, Gennaeodryas placens
- Western Yellow Robin, Eopsaltria griseogularis
- Eastern Yellow Robin, Eopsaltria australis
- White-breasted Robin, Eopsaltria georgiana
- White-faced Robin, Eopsaltria leucops
- Pale-yellow Robin, Eopsaltria capito
- Hooded Robin, Melanodryas cucullata
- Dusky Robin, Melanodryas vittata
- Mangrove Robin, Peneoenanthe pulverulenta
- White-rumped Robin, Peneothello bimaculata
- White-winged Robin, Peneothello sigillata
- Smoky Robin, Peneothello cryptoleuca
- Slaty Robin, Peneothello cyanus
Picathartoidea
The Picathartoidea are the next group to branch off separately from the Passerida tree, and did they ever branch! Somewhere in the gap between the Petroicoidea and Picathartoidea the passerids made the leap out of Australasia all the way to Africa. Which way they went, we are not sure. They may have come across the now-submerged Kerguelen Plateau. The Picathartoidea (rockfowl and rockjumpers) are both African endemics. They were considered Incertae sedis by Sibley and Monroe (1993) and there has been some discussion about whether they are best considered a corvid relative or part of Passerida. Several recent papers have made the case for placing them here (Barker et al., 2004; Beresford et al., 2005; Cracraft et al., 2004; Irestedt and Ohlson, 2008). Of course, once we have placed the Petroicidae in the Passerida, the choice is forced, Picathartoidea must also be in the Passerida. Recent research by Jønsson et al. (2007) found that the Rail-babbler belongs in the same clade. Each is distinct enough, and the genetic separation large enough, that we consider each a separate family in the Picathartoidea.
Picathartidae: Rockfowl Lowe, 1938
1 genus, 2 species HBW-12
- White-necked Rockfowl, Picathartes gymnocephalus
- Gray-necked Rockfowl, Picathartes oreas
Chaetopidae: Rockjumpers Fjeldså et al. 2015
1 genus, 2 species Not HBW Family
- Cape Rockjumper, Chaetops frenatus
- Orange-breasted Rockjumper / Drakensberg Rockjumper, Chaetops aurantius
Eupetidae: Rail-babbler Bonaparte, 1850
1 genus, 1 species HBW-12
- Rail-babbler, Eupetes macrocerus