STRISORES Baird, 1858
I've switched to using the older term Strisores (Baird 1858) for this group in place of Huxley's Cypselomorphae (1867). This allows the use of Cypselomorphae for a smaller clade, as in Mayr (2010).
Strisores were previously considered the biggest chunk of Metaves. The breakup of Metaves has made them the biggest chunk of Otidae. Although it is tempting to group all of the remaining nightbird families in a single order, it is hard to find convincing evidence that they are even constitute a clade (e.g., Barrowclough et al., 2006; Braun and Huddleston (2009), Mariaux and Braun, 1996; Mayr, 2002). The divisions between them are very deep, and DNA studies have not only not come to a clear conclusion about the 4 orders are related. Until Kimball et al. (2013), the evidence that Strisores formed a clade was a little shaky. Because of this, I have listed them in separate orders. Kimball et al. (2013), Hackett et al. (2008), and Ericson et al. (2006a) are three of genetic analyses that place them all with Apodiformes in Strisores. The Apodiformes may be found on the next page, with the remainder of the Strisores on this one.
Until Kimball et al. (2013), the monophyly of the Strisores was only supported by analyses that include β-fibrinogen. When β-fibrinogen was excluded in Ericson et al. (2006a), the Strisores ended up in the Anomalogonatae. Moreover, the nighthawks did not group with the other Strisores, being closer to a clade containing the owls and others. One of Braun and Huddleston (2009) analyses also placed the nighthawks closer to the owls. Kimball et al. (2013) included many more genes, and even if β-fibrinogen, there is still strong support for Strisores.
The orders are arranged as in Prum et al. (2015), who also estimate that each of the 5 orders dates back to the Palaeocene. My confidence in their topology is not high. Nonetheless, I think it is the best we currently have. Various other analyses have come to various conclusions. Hackett et al. (2008) have a different arrangement, with the Caprimulgiformes sister to the Apodiformes. Jarvis et al. (2014) is not helpful as they do not have sufficient coverage of these taxa. Recent morphological studies (e.g. Mayr, 2010; Nesbitt et al., 2011) suggest that the Steatornithiformes and Podargiformes are either basal branches or a sister clade to the remaining 3 orders. Sigurdsson (2013), using a combined molecular/morphological dataset, found that the Steatornithiformes were basal, with Nyctibiiformes and Caprimulgiformes as sisters, as were Podargiformes and Apodiformes.
CAPRIMULGIFORMES Ridgway, 1881
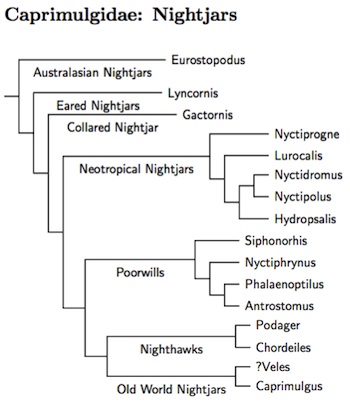
The thesis by Han (2006) and the follow-up paper (Han et al., 2010) have done much to clarify the situation within the Caprimulgidae. Sigurdsson's (2013) disseration and the following paper (Sigurdsson and Cracraft, 2014) touched up the overall structure a bit, and filled in more of the details (hence the species tree).
 |
| Click for species-level tree of Caprimulgidae |
|---|
Caprimulgidae: Nightjars, Nighthawks Vigors, 1825
16 genera, 100 species HBW-5
It was clear from previous work (Mariaux and Braun, 1996; Barrowclough et al. 2006; Larsen et al. 2007) that Caprimulgus itself was paraphyletic and that the Caprimulgidae were not simply divided into Nighthawks and Nightjars. It also seemed that the Eared-Nightjars are completely separate from Caprimulgidae. Both Han and Sigurdsson sampled many more taxa, and have established the general shape of the family, and shown that it doesn't make sense to separate some species into a separate family Eurostopodiae. Indeed, looking at both Han's and Sigurdsson's results, it does not appear that the wider Caprimulgidae can even be usefully divided in subfamilies. Such a division would not be particularly informative.
Han and Sigurdsson concur that the Great Eared-Nightjar, formerly placed in Eurostopdus is actually closer to the rest of Caprimulgidae. The Malaysian Eared-Nightjar is morphologically similar, and we are guessing that it is closely related to the Great Eared-Nightjar. Both are moved to the genus Lyncornis. The Papuan Nightjar has sometimes been placed in Lyncornis, but it is more closely related to the other Eurostopodus. Since it's thought to be close to the Cloud-forest and Satanic Nightjars, I leave them in Eurostopodus too.
The genus Eurostopodus has been enlarged a bit. The Solomons Nightjar, Eurostopodus nigripennis, has been split from the White-throated Nightjar, Eurostopodus mystacalis, and the New Caledonian Nightjar, Eurostopodus exul has been added based on a specimen collected in 1939. Both are discussed in Cleere (2010).
The next surprise is the Collared Nightjar, formerly Caprimulgus enarratus, which seems to be sister to the remaining nightjars and nighthawks. Han et al. (2010) established the new genus Gactornis for it. Sigurdsson (2013) agrees that it is the next branch of the Caprimulgidae.
The remaining Caprimulgidae seem to fall into 4 groups, a group of a group of Neotropical nighthawks and nightjars, an American group containing the poorwills and paruaques, the Chordeiles nighthawk group, and a complex group of Old World nightjars. Han (2006) had previously examined 3 genes, while Barrowclough et al. (2006) examined another (RAG-1). Sigurdsson (2013) included 4 genes, two of them novel. It seems clear enough that the Chordeiles nighthawks and the Old World nigthjars are sister groups. The relative arrangement of the other two with the Neotropical nightjars first, followed by the poorwills, and the the nighthawks and Old World nightjars seems to be the best bet, but a lack of consensus among the various genes.
The first group contains Nyctiprogne through Hydropsalis. I had been using different generic limits while the AOU's South American Checklist Committee deliberated on how to best incorporate Han et al. (2010). See SACC proposals #465, #501, and #522. Now that they have decided, the Sigurdsson dissertation comes along and emphasizes that this arrangement renders Setopagis paraphyletic.
The next best option is to submerge Setopagis, Eleothreptus, Systellura, and Macropsalis into Hydropsalis. That has been done here.
The group starts out with two basal nighthawk genera, Nyctiprogne and Lurocalis. This is followed by an expanded Nyctidromus (gaining the Scrub Nightjar from Caprimulgus) together with Nyctipolus (Ridgway 1912, type nigrescens), which includes the Blackish and Pygmy Nightjars. The Cayenne Nightjar, Nyctipolus maculosus, is only known from the type specimen. Cleere (2010) speculates that it is closely related to the Blackish Nightjar, so I have included it in Nyctipolus.
The remaining clade is now all in Hydropsalis. This clade has been augmented by several splits from the Band-winged Nightjar, Hydropsalis longirostris. They are marked with star's on the tree and include Tschudi's Nightjar, Hydropsalis decussata, and Tepui Nightjar, Hydropsalis roraimae, both also advocated by Cleere (2010), and Rufous-naped Nightjar, Hydropsalis ruficervix.
The second group consists of Siphonorhis, Nyctiphrynus, Phalaenoptilus, and the possibly unfamiliar genus Antrostomus. This name, due to Bonaparte (1838, type species A. carolinensis), is being applied to one of the New World Caprimulgus nightjars clades. As you can see, it is rather separated from the main group of Caprimulgus.
I have split the Whip-poor-will into Mexican Whip-poor-will, A. arizonae, and Eastern Whip-poor-will, A. vociferus. The Puerto Rican Nightjar (sometimes called Puerto Rican Whip-poor-will) has not been genetically tested. Han's (2006) analysis found the Mexican Whip-poor-will closer to the Dusky Nightjar than to the Eastern Whip-poor-will. However, Sigurdsson (2013) disagreed. The two Whip-poor-will's are vocally distinctive and differ somewhat in plumage, supporting their status as separate species (see also Howell and Webb, 1995; Navarro-Sigüenza and Peterson, 2004).
The third group consists of the Chordeiles nighthawks and the Nacunda Nighthawk, Podager nacunda. Rather than follow the AOU's SACC in placing them in Chordeiles I now follow the alternative of putting the Least Nighthawk in Podager.
None of the papers have densely sampled the Old World nightjars. I've put the Brown Nightjar, Veles binotatus, in group four on the grounds that is a Old World species sometimes considered part of Caprimulgus. However, others have put it elsewhere (e.g., closer to Chordeiles or Lyncornis), which is also consistent with the linear order. Han (2006) found considerable structure in the remaining Caprimulgus. As a general rule, the Asian species appear to be relatively basal in Caprimulgus. This is not reflected in the current arrangement of the species, which is fairly conventional. Note that Macrodipteryx needed to be submerged into Caprimulgus.
Finally, ABA and AOU use the name Gray Nightjar for Caprimulgus indicus. Here C. jotaka has been split from C. indicus and keeps the name Gray Nightjar while C. indicus becomes Jungle Nightjar. The third subspecies of the former Gray Nightjar is also split as Palau Nightjar, Caprimulgus phalaena (Cleere, 2010).
- Spotted Nightjar, Eurostopodus argus
- White-throated Nightjar, Eurostopodus mystacalis
- Solomons Nightjar, Eurostopodus nigripennis
- New Caledonian Nightjar, Eurostopodus exul
- Satanic Nightjar, Eurostopodus diabolicus
- Papuan Nightjar, Eurostopodus papuensis
- Archbold's Nightjar, Eurostopodus archboldi
- Malaysian Eared-Nightjar, Lyncornis temminckii
- Great Eared-Nightjar, Lyncornis macrotis
- Collared Nightjar, Gactornis enarratus
- Band-tailed Nighthawk, Nyctiprogne leucopyga
- Bahian Nighthawk / Plain-tailed Nighthawk, Nyctiprogne vielliardi
- Short-tailed Nighthawk, Lurocalis semitorquatus
- Rufous-bellied Nighthawk, Lurocalis rufiventris
- Common Pauraque / Pauraque, Nyctidromus albicollis
- Scrub Nightjar / Anthony's Nightjar, Nyctidromus anthonyi
- Blackish Nightjar, Nyctipolus nigrescens
- Cayenne Nightjar, Nyctipolus maculosus
- Pygmy Nightjar, Nyctipolus hirundinaceus
- Roraiman Nightjar, Hydropsalis whitelyi
- Swallow-tailed Nightjar, Hydropsalis segmentata
- Lyre-tailed Nightjar, Hydropsalis lyra
- Tschudi's Nightjar, Hydropsalis decussata
- Todd's Nightjar, Hydropsalis heterura
- Little Nightjar, Hydropsalis parvula
- White-winged Nightjar, Hydropsalis candicans
- Sickle-winged Nightjar, Hydropsalis anomala
- Band-winged Nightjar, Hydropsalis longirostris
- Rufous-naped Nightjar, Hydropsalis ruficervix
- Tepui Nightjar, Hydropsalis roraimae
- Spot-tailed Nightjar, Hydropsalis maculicaudus
- Long-trained Nightjar, Hydropsalis forcipata
- White-tailed Nightjar, Hydropsalis cayennensis
- Ladder-tailed Nightjar, Hydropsalis climacocerca
- Scissor-tailed Nightjar, Hydropsalis torquata
- Jamaican Pauraque / Jamaican Poorwill, Siphonorhis americana
- Least Pauraque / Least Poorwill, Siphonorhis brewsteri
- Choco Poorwill, Nyctiphrynus rosenbergi
- Eared Poorwill, Nyctiphrynus mcleodii
- Yucatan Poorwill, Nyctiphrynus yucatanicus
- Ocellated Poorwill, Nyctiphrynus ocellatus
- Common Poorwill, Phalaenoptilus nuttallii
- Tawny-collared Nightjar, Antrostomus salvini
- Buff-collared Nightjar, Antrostomus ridgwayi
- Dusky Nightjar, Antrostomus saturatus
- Mexican Whip-poor-will, Antrostomus arizonae
- Eastern Whip-poor-will, Antrostomus vociferus
- Puerto Rican Nightjar, Antrostomus noctitherus
- Yucatan Nightjar, Antrostomus badius
- Silky-tailed Nightjar, Antrostomus sericocaudatus
- Rufous Nightjar, Antrostomus rufus
- Chuck-will's-widow, Antrostomus carolinensis
- Greater Antillean Nightjar, Antrostomus cubanensis
- Nacunda Nighthawk, Podager nacunda
- Least Nighthawk, Podager pusillus
- Lesser Nighthawk, Chordeiles acutipennis
- Sand-colored Nighthawk, Chordeiles rupestris
- Common Nighthawk, Chordeiles minor
- Antillean Nighthawk, Chordeiles gundlachii
- Brown Nightjar, Veles binotatus
- Red-necked Nightjar, Caprimulgus ruficollis
- Jungle Nightjar, Caprimulgus indicus
- Gray Nightjar, Caprimulgus jotaka
- Palau Nightjar, Caprimulgus phalaena
- European Nightjar, Caprimulgus europaeus
- Sombre Nightjar, Caprimulgus fraenatus
- Rufous-cheeked Nightjar, Caprimulgus rufigena
- Egyptian Nightjar, Caprimulgus aegyptius
- Sykes's Nightjar, Caprimulgus mahrattensis
- Vaurie's Nightjar, Caprimulgus centralasicus
- Nubian Nightjar, Caprimulgus nubicus
- Golden Nightjar, Caprimulgus eximius
- Jerdon's Nightjar, Caprimulgus atripennis
- Large-tailed Nightjar, Caprimulgus macrurus
- Mees's Nightjar, Caprimulgus meesi
- Andaman Nightjar, Caprimulgus andamanicus
- Philippine Nightjar, Caprimulgus manillensis
- Sulawesi Nightjar, Caprimulgus celebensis
- Donaldson Smith's Nightjar, Caprimulgus donaldsoni
- Black-shouldered Nightjar, Caprimulgus nigriscapularis
- Fiery-necked Nightjar, Caprimulgus pectoralis
- Montane Nightjar, Caprimulgus poliocephalus
- Ruwenzori Nightjar, Caprimulgus ruwenzorii
- Indian Nightjar, Caprimulgus asiaticus
- Madagascan Nightjar, Caprimulgus madagascariensis
- Swamp Nightjar, Caprimulgus natalensis
- Nechisar Nightjar, Caprimulgus solala
- Plain Nightjar, Caprimulgus inornatus
- Star-spotted Nightjar, Caprimulgus stellatus
- Savanna Nightjar, Caprimulgus affinis
- Freckled Nightjar, Caprimulgus tristigma
- Bonaparte's Nightjar, Caprimulgus concretus
- Salvadori's Nightjar, Caprimulgus pulchellus
- Prigogine's Nightjar, Caprimulgus prigoginei
- Bates's Nightjar, Caprimulgus batesi
- Long-tailed Nightjar, Caprimulgus climacurus
- Slender-tailed Nightjar, Caprimulgus clarus
- Square-tailed Nightjar, Caprimulgus fossii
- Standard-winged Nightjar, Caprimulgus longipennis
- Pennant-winged Nightjar, Caprimulgus vexillarius
STEATORNITHIFORMES Sharpe 1891
The results of both Hackett et al. (2008) and Braun and Huddleston (2009) suggest the oilbirds and potoos are sisters, which makes sense geographically. Note however, that oilbirds were once more widely distributed. An oilbird fossil is known from Wyoming (Olson, 1987).
Steatornithidae: Oilbird Bonaparte, 1842
1 genus, 1 species HBW-5
- Oilbird, Steatornis caripensis
NYCTIBIIFORMES Informal
No one seems to have recongized the Nyctibiiformes at an ordinal level before. I've been using it on these webpages since at least March 2008. If had I published it in print instead of a web page I could have my name on it!
Nyctibiidae: Potoos Chenu & des Murs, 1851
1 genus, 7 species HBW-5
- Rufous Potoo, Nyctibius bracteatus
- Great Potoo, Nyctibius grandis
- Long-tailed Potoo, Nyctibius aethereus
- Common Potoo, Nyctibius griseus
- Northern Potoo, Nyctibius jamaicensis
- Andean Potoo, Nyctibius maculosus
- White-winged Potoo, Nyctibius leucopterus
PODARGIFORMES Sharpe 1891
The Frogmouths are sometimes divided into two families, Podargidae and Batrachostomidae. Molecular evidence indicates they are each other's closest relatives, and I think that is best indicated by placing them in a single family. One new species, the Solomon Islands Frogmouth, was recently discovered (Cleere et al., 2007). It seems to be closer to Podargus than to Batrachostomus.
Following Cleere (2010), Blyth's Frogmouth, Batrachostomus affinis and Palawan Frogmouth, Batrachostomus chaseni) are separated from Javan Frogmouth, Batrachostomus javensis, based on differences in vocalizations. Although Cleere separates it as a separate species, there seems to be some question concerning the vocalizations of the race continentalis (see Marshall, 1978). Until that is clarified, preferably in a journal article, I continue to include it as a subspecies of Blyth's Frogmouth, Batrachostomus affinis.
Although the extant frogmouths are restricted to the Oriental and Australasian regions, they were not always so. Frogmouth fossils have been found in both Europe and the United States (Nesbitt et al., 2011). The American fossils (Fluvioviridavis) are from the Fossil Butte Member of the Green River Formation, and date to approximately 52 million years ago (Grande, 2013). This age underscores the fact that the various orders in the Strisores represent deep divisions in the avian tree.
Nesbitt et al. (2011) also note: “Intriguingly, FBM avian taxa evaluated phylogenetically to date have overwhelmingly been placed along the stem lineages of extant avian subclades comprising traditional `orders' or `families'.”
Podargidae: Frogmouths G.R. Gray, 1847
3 genera, 16 species HBW-5
- Solomons Frogmouth, Rigidipenna inexpectata
- Marbled Frogmouth, Podargus ocellatus
- Papuan Frogmouth, Podargus papuensis
- Tawny Frogmouth, Podargus strigoides
- Large Frogmouth, Batrachostomus auritus
- Dulit Frogmouth, Batrachostomus harterti
- Philippine Frogmouth, Batrachostomus septimus
- Gould's Frogmouth, Batrachostomus stellatus
- Sri Lanka Frogmouth, Batrachostomus moniliger
- Hodgson's Frogmouth, Batrachostomus hodgsoni
- Short-tailed Frogmouth, Batrachostomus poliolophus
- Bornean Frogmouth, Batrachostomus mixtus
- Javan Frogmouth, Batrachostomus javensis
- Blyth's Frogmouth, Batrachostomus affinis
- Palawan Frogmouth, Batrachostomus chaseni
- Sunda Frogmouth, Batrachostomus cornutus