NEOAVES Sibley, Ahlquist, & Monroe, 2008
Neoaves is the sister clade to Galloanserae. Jarvis et al. (2014) show that it has two parts: Columbea and Passerea.
COLUMBEA Jarvis et al., 2014
Columbea contains about 3½% of the avian tree. The sister relationship between Columbimorphae and Mirandornithes in Jarvis et al. (2014) is not so suprising as they have been included in Metaves since it was proposed by Fain and Houde (2004).
Jarvis et al.'s results make it clear that the original Metaves hypothesis does not hold up in larger datasets. It was in fact an artifact of using the β-fibrogen gene, particularly the 7th intron. Nonetheless, some of the signal was real, and Columbea remains once the Eurypygimorphae, Strisores, and Hoatzin have been moved elsewhere.
Mirandornithes Sangster, 2005
Van Tuinen et al. (2001) seem to have been the first to recognize that flamingos and grebes were sister taxa. Their reanalysis of Sibley and Ahlquist (1990) shows that other data to support this association were available earlier, but that the association had been missed by Sibley and Ahlquist, who did not include both of them in any of their individual trees. They did not suspect flamingos and grebes could be closely related.
Later analysis — Chubb (2004), Cracraft et al. (2004), Ericson et al. (2006a), Brown et al. (2008), Hackett (2008), Gibb et al. (2013), McCormack et al. (2013), and Jarvis et al. (2014) — have also supported Mirandornithes. Further, Mayr (2004) identified morphological and oological evidence in favor of Mirandornithes. See also Mayr (2007, 2008). As with the genetic data, previous morphological investigations also did not consider the possibility of a relationship here. It was just too inconceivable.
It's optional whether to lump the grebes and flamingos into one order, or treat them as two. I've switched to the conventional treatment with two orders. The large morphological differences and ancient split from the flamingos (e.g., Jarvis et al., 2014; Brown et al., 2008) support this.
PHOENICOPTERIFORMES Fürbringer, 1888
Phoenicopteridae: Flamingos Bonaparte, 1831

The genetics of flamingos were studied by Torres et al. (2014). They found that all 6 flamingos shared a recent common ancestor, perhaps 4-6 million years ago. They also recommended sinking Phoeniconaias into Phoenicoparrus, as they considered the distinction between them “arbitrary and uninformative”. Based on the fossil record, Torres et al. (2014) also suggested that the grebe-flamingo split occurred around the beginning of the Oligocene, i.e., about 33 million years ago. Mayr (2014a), using morphological evidence, found that the Eocene Juncitarus is a stem representative of Mirandornithes. As such, it gives little information about the timing of the flamingo-grebe, although Mayr suggested middle Eocene. Jarvis et al. (2014) date the split at about 56 mya, roughly at the beginning of the Eocene. I have to wonder whether the crown group may also not be as recent as Torres et al. think, especially since they suggest that their divergence dates among flamingos fit better with an even more recent split between grebes and flamingos. This suggests to me that the older mitochondrial dates are more likely to be correct, and that the true date is probably toward (or past!) the long end of the range. The SACC has decided to keep the current genera, and I follow them here.
3 genera, 6 species HBW-1
- Chilean Flamingo, Phoenicopterus chilensis
- Greater Flamingo, Phoenicopterus roseus
- American Flamingo, Phoenicopterus ruber
- Lesser Flamingo, Phoeniconaias minor
- Andean Flamingo, Phoenicoparrus andinus
- James's Flamingo, Phoenicoparrus jamesi
PODICIPEDIFORMES Fürbringer, 1888
Podicipedidae: Grebes Bonaparte, 1831
6 genera, 24 species HBW-1
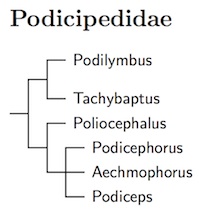
There's no complete molecular phylogeny of the grebes. However, Fjeldså (2004) provides a morphological phylogeny. Christidis and Boles (2008) have also adopted Fjeldså's phylogeny. For the present, I also use it with some modification based on Ogawa et al. (2015).
Ogawa et al. (2015) found that Rollandia is embedded in Podiceps, so it has been merged into Podiceps. The arrangement within the expanded Podiceps is based on Ogawa et al. (2015).
Bochenski's (1994) osteological study of the grebes found that the Great Grebe is significantly different from the other Podiceps grebes. He created the genus Podicephorus for it. Fjeldså endorsed this view by putting Podicephorus in an unresolved trichotomy with Aechmophorus and the remaining Podiceps. The SACC has yet to consider the issue.
Tricolored Grebe, Tachybaptus tricolor, has been split from Little Grebe, Tachybaptus ruficollis. The subspecies vulcanorum is included in Tricolored Grebe. See Mlíkovský (2010).
Based on Ogawa et al. (2015), the Eared Grebe, Podiceps californicus, has been split from the Black-necked Grebe, Podiceps nigricollis. Ogawa et al. did not find a clear separation between Silvery Grebe, Podiceps occipitalis, and Junin Grebe, Podiceps taczanowskii. Further study is needed to clarify the situation.
- Pied-billed Grebe, Podilymbus podiceps
- Atitlan Grebe, Podilymbus gigas
- Alaotra Grebe, Tachybaptus rufolavatus
- Little Grebe, Tachybaptus ruficollis
- Tricolored Grebe, Tachybaptus tricolor
- Australasian Grebe, Tachybaptus novaehollandiae
- Madagascan Grebe, Tachybaptus pelzelnii
- Least Grebe, Tachybaptus dominicus
- Hoary-headed Grebe, Poliocephalus poliocephalus
- New Zealand Grebe, Poliocephalus rufopectus
- Great Grebe, Podicephorus major
- Western Grebe, Aechmophorus occidentalis
- Clark's Grebe, Aechmophorus clarkii
- Red-necked Grebe, Podiceps grisegena
- Great Crested Grebe, Podiceps cristatus
- Horned Grebe / Slavonian Grebe, Podiceps auritus
- White-tufted Grebe, Podiceps rolland
- Titicaca Grebe, Podiceps micropterus
- Hooded Grebe, Podiceps gallardoi
- Silvery Grebe, Podiceps occipitalis
- Junin Grebe, Podiceps taczanowskii
- Black-necked Grebe, Podiceps nigricollis
- Eared Grebe, Podiceps californicus
- Colombian Grebe, Podiceps andinus
Columbimorphae Latham, 1790
MESITORNITHIFORMES Wetmore 1960
Sharpe had earlier tried to establish such a suborder (Mesitides), but based it on the preoccupied genus name Mesites. I haven't been able to find uses of Mesitornis as a type genus earlier than Wetmore.
Mesitornithidae: Mesites Wetmore, 1960 (1850)
2 genera, 3 species HBW-3
- Subdesert Mesite, Monias benschi
- White-breasted Mesite, Mesitornis variegatus
- Brown Mesite, Mesitornis unicolor
PTEROCLIFORMES Huxley, 1868
Huxley (1868) first separated the sandgrouse as an order, using the name Pteroclomorphae, which modernizes to Pterocliformes.
I follow Jarvis et al. (2014) where the mesites are sister to the sandgrouse, with the combination sister to the doves and pigeons. Hackett et al. (2008) also considered the three groups a clade, but with a different arrangement. Jarvis et al. also estimated that the divisions between the three clades date back to the early Paleocene.
Pteroclidae: Sandgrouse Bonaparte, 1831
2 genera, 16 species HBW-4
 |
| Click for species-level Pteroclidae tree |
|---|
There has been some controversy about how to spell the family name. Although TiF has used Pteroclidae for a long time, back in 2010 both Pteroclidae (Clements, HBW, Sibley-Monroe) and Pteroclididae (AOU, BLI, Howard-Moore, IOC) were in general use, and even Pterocleidae had also been used. Since then, Pteroclidae has won the day. The name indicates it is known for its wing, i.e., “-cles” takes the same meaning as in names such as Heracles. By analogy with Heraclidae/Heracleidae, it would then appear that either Pteroclidae or Pterocleidae would be correct. Most importantly, the first is the form used by Bonaparte when he established the family-group name in 1831 (as the subfamily Pteroclinae), and is used here.
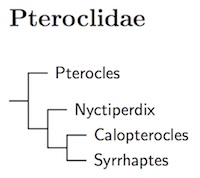
The arrangement here is based on Cohen (2011). There's a bit of ambiguity in Cohen's results. According to the nuclear DNA, the Pin-tailed Sandgrouse, Pterocles alchata, is the basal species. However, mitochondrial DNA puts it sister to Burchell's Sandgrouse, Calopterocles burchelli, albeit with mediocre support. Because of that ambiguity, I've put Burchell's Sandgrouse in its own genus, Calopterocles (Roberts, 1922). Further, Pterocles is now restricted to the Pin-tailed Sandgrouse. Most of the wedge-tailed sandgrouse, with a long 10th primary have been of the former Pterocles species have been moved to Syrrhaptes, which is buried deep inside that clade. The other former Pterocles have been separated as Nyctiperdix (Roberts, 1922, type bicinctus).
- Pin-tailed Sandgrouse, Pterocles alchata
- Black-faced Sandgrouse, Nyctiperdix decoratus
- Double-banded Sandgrouse, Nyctiperdix bicinctus
- Four-banded Sandgrouse, Nyctiperdix quadricinctus
- Lichtenstein's Sandgrouse, Nyctiperdix lichtensteinii
- Painted Sandgrouse, Nyctiperdix indicus
- Burchell's Sandgrouse, Calopterocles burchelli
- Yellow-throated Sandgrouse, Syrrhaptes gutturalis
- Crowned Sandgrouse, Syrrhaptes coronatus
- Madagascan Sandgrouse, Syrrhaptes personatus
- Spotted Sandgrouse, Syrrhaptes senegallus
- Chestnut-bellied Sandgrouse, Syrrhaptes exustus
- Namaqua Sandgrouse, Syrrhaptes namaqua
- Black-bellied Sandgrouse, Syrrhaptes orientalis
- Tibetan Sandgrouse, Syrrhaptes tibetanus
- Pallas's Sandgrouse, Syrrhaptes paradoxus
COLUMBIFORMES Latham, 1790
Columbidae: Doves, Pigeons Leach, 1820
47 genera, 341 species HBW-4
 |
| Click for genus-level Columbidae tree |
|---|
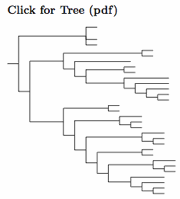
I have based the organization of the Columbidae on the paper by Pereira et al. (2007), which provides a comprehensive DNA-based phylogentic tree. Although the dodos and Rodriguez Solitaire (genera Raphus and Pezophaps) have been traditionally considered a separate family in the Columbiformes, the DNA says otherwise. Shapiro et al. (2002) and Pereira et al. (2007) found that these two genera are buried deeply within the Columbidae, in the Raphinae. The Reunion Solitaire seems to have actually been an ibis! See Mourer-Chauviré et al. (1995).
The DNA testing shows three major clades. A basal clade includes a subclade consisting of the New World genera Geotrygon, Leptotila, Zenaida, together with a subclade of New World pigeons (including the Passenger Pigeon), as well as typical pigeons, cuckoo-doves, and turtle-doves. It is sister to the other two clades together. One of them consists of the New World Ground-Doves. The other contains all other doves. I rank the three major clades as subfamilies: Columbinae, Claravinae, and Raphinae.
The name Peristerinae is sometimes used for Claravinae. However, this is incorrect as Peristerinae is based on the genus Peristera (Swainson 1827), which is a junior homonym of the mollusc genus Peristera (Rafinesque 1815). Thus Peristerinae is not available. The genus Peristera (Swainson 1827) was replaced by Claravis (Oberholser 1899). Todd (not Richmond) established the subfamily Claravinae in 1917, which he misspelled as Claraviinae (Dickinson and Raty, 2015).
The arrangement of Claravinae is based on Sweet and Johnson (2015). As a result, the Purple-winged Ground-Dove, Claravis geoffroyi and Maroon-chested Ground-Dove, Claravis mondetoura have been moved to Metriopelia.
The name Raphidae has often been used for a family containing dodos and solitaires. However, the dodos and solitaires turn out to be nested well within our subfamily Raphinae, and even deeper in the family Columbidae.
The subfamily Columbinae has been studied in more detail by Johnson and Clayton (2000), Johnson et al. (2001), Gonzalez et al. (2009a), and Johnson and Weckstein (2011). Zenaidini and the species in Streptopelia and Columba, but not Patagioenas, have been rearranged accordingly. Although Johnson et al. (2001) argued that Nesoenas should be merged into Streptopelia, Cheke (2005) makes the case for instead moving the Malagasy Turtle-Dove to Nesoenas and distinguishing the Laughing and Spotted Doves in a separate genus. The results of Gonalez et al. (2009a) support this move.
In the case of Nesoenas, both Nesoenas and Homopelia (type picturatus) have equal priority, being both named in the same work of Salvadori 1893. Cheke acts as first reviser, choosing Nesoenas. The Laughing Dove is the type of Stigmatopelia (Sundevall 1873) while the Spotted Dove is the type of Spilopelia (also Sundevall 1873). Cheke attempts to designate Stigmatopelia as the genus, but as John Penhallurick points out Schodde and Mason had previously (1997) chosen Spilopelia, so Spilopelia they become.
The work by Johnson and Weckstein (2011) showed that Geotrygon as usually constituted is paraphyletic with its members belonging to 3 separate clades. The Olive-backed Quail-Dove is sister to Leptotila. Banks et al. (2013) establish the genus Leptotrygon for it. The remaining quail-doves fall into at least two groups. Although Johnson and Weckstein did not include the type of Geotrygon (versicolor), a further analysis by Banks et al. (2013) included it and chrysia. The other “Geotrygon” form a clade sister to Zenaida. Banks et al. (2013) have now established the name Zentrygon for it (type costaricensis).
The English names of White-faced Dove, Turacoena manadensis, and Black Dove, Turacoena modesta, have been changed to White-faced Cuckoo-Dove and Black Cuckoo-Dove to match recent IOC changes. There are also several splits.
- Based on Ng and Rheindt (2016), Sula Cuckoo-Dove, Turacoena sulaensis, has been split from White-faced Cuckoo-Dove, Turacoena manadensis.
The remaining splits are based on Ng et al. (2016).
- Slender-billed Cuckoo-Dove, Macropygia amboinensis, has been split into Sultan's Cuckoo-Dove, Macropygia doreya, and Amboyna Cuckoo-Dove, Macropygia amboinensis.
- Bar-necked Cuckoo-Dove, Macropygia magna, has been split into Flores Sea Cuckoo-Dove, Macropygia macassariensis, Timor Cuckoo-Dove, Macropygia magna, and Tanimbar Cuckoo-Dove, Macropygia timorlaoensis.
- Enggano Cuckoo-Dove, Macropygia cinnamomea, and Barusan Cuckoo Dove, Macropygia modiglianii, have been split from Ruddy Cuckoo-Dove, Macropygia emiliana.
- The subspecies borneensis has been moved from Ruddy Cuckoo-Dove, Macropygia emiliana, to Philippine Cuckoo-Dove, Macropygia tenuirostris.
Jønsson et al. (2011a) and Moyle et al. (2013) studied the Alopecoenas ground-doves and Gallicolumba bleeding-hearts. Jønsson et al. found that Gallicolumba was paraphyletic, and recommended splitting it into Gallicolumba (bleeding-hearts) and Alopecoenas (Australasian ground-doves). Their phylogenetic trees seemed to indicate there was still paraphyly even with the restricted Gallicolumba. The further analysis by Moyle et al. revealed issues with pseudogenes that were causing some confusion. Their results support placing the Crested Pigeon in the genus Ocyphaps. Finally, the three extinct Alopecoenas are most likely closely related to A. sanctaecrucis and A. stairi, and their placement reflects this.
The Purple Quail-Dove / Indigo-crowned Quail-Dove Geotrygon purpurata, has been split from Sapphire Quail-Dove, Geotrygon saphirina based on SACC proposal #566.
Gibb and Penny (2010) investigated the fruit-doves and close relatives. They confirmed Shapiro et al.'s (2002) result that Alectroenas and Drepanoptila are embedded in fruit-dove genus Ptilinopus. Cibois et al. (2014) concured in their more complete analysis of the enlarged Ptilinopus. Moreover, they were able to include most Ptilinopus species. The current arrangement of Ptilinopus is based on their results. They suggested a six genus treatment that retained Alectroenas and Drepanoptila. However, this may mean that Ptilinopus is not monophyletic, and it does not materially solve the problem of Ptilinopus heterogeneity. I do not recommend it at this time.
The Black-banded Fruit-Dove, Ptilinopus alligator, has been split from Banded Fruit-Dove, Ptilinopus cinctus. Given present knowledge, it's fairly arbitrary whether to split or lump these allopatric forms. However, one is in Australia, the other in the Lesser Sundas, and the current tendency is for such forms to be split.
Based on Rheindt et al. (2011a), the Maroon-chinned Fruit-Dove, Ptilinopus subgularis is split into 3 species: Oberholser's Fruit-Dove, Ptilinopus epius, Banggai Fruit-Dove, Ptilinopus subgularis, and Sula Fruit-Dove, Ptilinopus mangoliensis.
Columbinae Leach, 1820
Zenaidini Bonaparte, 1853
- Blue-headed Quail-Dove, Starnoenas cyanocephala
- Purple Quail-Dove / Indigo-crowned Quail-Dove, Geotrygon purpurata
- Sapphire Quail-Dove, Geotrygon saphirina
- Crested Quail-Dove, Geotrygon versicolor
- Gray-fronted Quail-Dove, Geotrygon caniceps
- White-fronted Quail-Dove, Geotrygon leucometopia
- Ruddy Quail-Dove, Geotrygon montana
- Violaceous Quail-Dove, Geotrygon violacea
- Key West Quail-Dove, Geotrygon chrysia
- Bridled Quail-Dove, Geotrygon mystacea
- Olive-backed Quail-Dove, Leptotrygon veraguensis
- White-tipped Dove, Leptotila verreauxi
- Caribbean Dove, Leptotila jamaicensis
- Large-tailed Dove / Yungas Dove, Leptotila megalura
- Gray-fronted Dove, Leptotila rufaxilla
- Gray-chested Dove, Leptotila cassinii
- Tolima Dove, Leptotila conoveri
- Ochre-bellied Dove, Leptotila ochraceiventris
- Gray-headed Dove, Leptotila plumbeiceps
- Brown-backed Dove / Azuero Dove, Leptotila battyi
- Pallid Dove, Leptotila pallida
- Grenada Dove, Leptotila wellsi
- White-winged Dove, Zenaida asiatica
- West Peruvian Dove, Zenaida meloda
- Zenaida Dove, Zenaida aurita
- Galapagos Dove, Zenaida galapagoensis
- Eared Dove, Zenaida auriculata
- Mourning Dove, Zenaida macroura
- Socorro Dove, Zenaida graysoni
- Buff-fronted Quail-Dove, Zentrygon costaricensis
- Tuxtla Quail-Dove, Zentrygon carrikeri
- Purplish-backed Quail-Dove, Zentrygon lawrencii
- White-faced Quail-Dove, Zentrygon albifacies
- White-throated Quail-Dove, Zentrygon frenata
- Lined Quail-Dove, Zentrygon linearis
- Chiriqui Quail-Dove, Zentrygon chiriquensis
- Russet-crowned Quail-Dove, Zentrygon goldmani
Columbini Leach, 1820
- Great Cuckoo-Dove, Reinwardtoena reinwardti
- Pied Cuckoo-Dove, Reinwardtoena browni
- Crested Cuckoo-Dove, Reinwardtoena crassirostris
- Black Cuckoo-Dove, Turacoena modesta
- White-faced Cuckoo-Dove, Turacoena manadensis
- Sula Cuckoo-Dove, Turacoena sulaensis
- Barred Cuckoo-Dove, Macropygia unchall
- Andaman Cuckoo-Dove, Macropygia rufipennis
- Barusan Cuckoo-Dove, Macropygia modiglianii
- Philippine Cuckoo-Dove, Macropygia tenuirostris
- Enggano Cuckoo-Dove, Macropygia cinnamomea
- Ruddy Cuckoo-Dove, Macropygia emiliana
- Sultan's Cuckoo-Dove, Macropygia doreya
- Flores Sea Cuckoo-Dove, Macropygia macassariensis
- Amboyna Cuckoo-Dove, Macropygia amboinensis
- Timor Cuckoo-Dove, Macropygia magna
- Tanimbar Cuckoo-Dove, Macropygia timorlaoensis
- Brown Cuckoo-Dove, Macropygia phasianella
- Bar-tailed Cuckoo-Dove, Macropygia nigrirostris
- MacKinlay's Cuckoo-Dove, Macropygia mackinlayi
- Little Cuckoo-Dove, Macropygia ruficeps
- Passenger Pigeon, Ectopistes migratorius
- White-crowned Pigeon, Patagioenas leucocephala
- Scaly-naped Pigeon, Patagioenas squamosa
- Scaled Pigeon, Patagioenas speciosa
- Picazuro Pigeon, Patagioenas picazuro
- Bare-eyed Pigeon, Patagioenas corensis
- Spot-winged Pigeon, Patagioenas maculosa
- Band-tailed Pigeon, Patagioenas fasciata
- Chilean Pigeon, Patagioenas araucana
- Ring-tailed Pigeon, Patagioenas caribaea
- Pale-vented Pigeon, Patagioenas cayennensis
- Red-billed Pigeon, Patagioenas flavirostris
- Peruvian Pigeon / Maranon Pigeon, Patagioenas oenops
- Plain Pigeon, Patagioenas inornata
- Plumbeous Pigeon, Patagioenas plumbea
- Ruddy Pigeon, Patagioenas subvinacea
- Short-billed Pigeon, Patagioenas nigrirostris
- Dusky Pigeon, Patagioenas goodsoni
- Red Turtle-Dove, Streptopelia tranquebarica
- Island Collared-Dove, Streptopelia bitorquata
- Oriental Turtle-Dove, Streptopelia orientalis
- European Turtle-Dove, Streptopelia turtur
- Dusky Turtle-Dove, Streptopelia lugens
- Adamawa Turtle-Dove, Streptopelia hypopyrrha
- Ring-necked Dove, Streptopelia capicola
- Vinaceous Dove, Streptopelia vinacea
- Red-eyed Dove, Streptopelia semitorquata
- Mourning Collared-Dove, Streptopelia decipiens
- White-winged Collared-Dove, Streptopelia reichenowi
- Eurasian Collared-Dove, Streptopelia decaocto
- African Collared-Dove, Streptopelia roseogrisea
- Laughing Dove, Spilopelia senegalensis
- Spotted Dove, Spilopelia chinensis
- Malagasy Turtle-Dove, Nesoenas picturatus
- Pink Pigeon, Nesoenas mayeri
- Rodrigues Pigeon, Nesoenas rodericanus
- Lemon Dove, Columba larvata
- Eastern Bronze-naped Pigeon, Columba delegorguei
- Western Bronze-naped Pigeon, Columba iriditorques
- Island Bronze-naped Pigeon, Columba malherbii
- Ashy Woodpigeon, Columba pulchricollis
- Nilgiri Woodpigeon, Columba elphinstonii
- Sri Lanka Woodpigeon, Columba torringtoniae
- Pale-capped Pigeon, Columba punicea
- Silvery Pigeon, Columba argentina
- Andaman Woodpigeon, Columba palumboides
- Laurel Pigeon, Columba junoniae
- African Olive-Pigeon, Columba arquatrix
- Cameroon Olive-Pigeon, Columba sjostedti
- Sao Tome Olive-Pigeon, Columba thomensis
- Comoros Olive-Pigeon, Columba pollenii
- Speckled Woodpigeon, Columba hodgsonii
- White-naped Pigeon, Columba albinucha
- Common Woodpigeon, Columba palumbus
- Trocaz Pigeon, Columba trocaz
- Bolle's Pigeon, Columba bollii
- Afep Pigeon, Columba unicincta
- Speckled Pigeon, Columba guinea
- White-collared Pigeon, Columba albitorques
- Stock Dove, Columba oenas
- Yellow-eyed Pigeon, Columba eversmanni
- Somali Pigeon, Columba oliviae
- Rock Pigeon / Rock Dove, Columba livia
- Hill Pigeon, Columba rupestris
- Snow Pigeon, Columba leuconota
- Japanese Woodpigeon, Columba janthina
- Bonin Woodpigeon, Columba versicolor
- Ryukyu Woodpigeon, Columba jouyi
- Metallic Pigeon, Columba vitiensis
- White-headed Pigeon, Columba leucomela
- Yellow-legged Pigeon, Columba pallidiceps
Claravinae: American Ground-Doves Todd, 1917
- Blue Ground-Dove, Claravis pretiosa
- Long-tailed Ground-Dove, Uropelia campestris
- Maroon-chested Ground-Dove, Metriopelia mondetoura
- Purple-winged Ground-Dove, Metriopelia geoffroyi
- Golden-spotted Ground-Dove, Metriopelia aymara
- Black-winged Ground-Dove, Metriopelia melanoptera
- Bare-faced Ground-Dove, Metriopelia ceciliae
- Bare-eyed Ground-Dove / Moreno's Ground-Dove, Metriopelia morenoi
- Blue-eyed Ground-Dove, Columbina cyanopis
- Picui Ground-Dove, Columbina picui
- Croaking Ground-Dove, Columbina cruziana
- Inca Dove, Columbina inca
- Scaled Dove, Columbina squammata
- Common Ground-Dove, Columbina passerina
- Plain-breasted Ground-Dove, Columbina minuta
- Ruddy Ground-Dove, Columbina talpacoti
- Ecuadorian Ground-Dove, Columbina buckleyi
Raphinae: Old World Doves and Pigeons Wetmore, 1930 (1835)
Phabini: Australasian Pigeons and Doves Bonaparte, 1853
- New Guinea Bronzewing, Henicophaps albifrons
- New Britain Bronzewing, Henicophaps foersteri
- Sulawesi Ground-Dove, Gallicolumba tristigmata
- Cinnamon Ground-Dove, Gallicolumba rufigula
- Sulu Bleeding-heart, Gallicolumba menagei
- Mindoro Bleeding-heart, Gallicolumba platenae
- Negros Bleeding-heart, Gallicolumba keayi
- Mindanao Bleeding-heart, Gallicolumba crinigera
- Luzon Bleeding-heart, Gallicolumba luzonica
- Wetar Ground-Dove, Alopecoenas hoedtii
- White-breasted Ground-Dove, Alopecoenas jobiensis
- Marquesan Ground-Dove, Alopecoenas rubescens
- White-fronted Ground-Dove, Alopecoenas kubaryi
- Polynesian Ground-Dove, Alopecoenas erythroptera
- White-throated Ground-Dove, Alopecoenas xanthonura
- Bronze Ground-Dove, Alopecoenas beccarii
- Palau Ground-Dove, Alopecoenas canifrons
- Thick-billed Ground-Dove, Alopecoenas salamonis
- Santa Cruz Ground-Dove, Alopecoenas sanctaecrucis
- Tongan Ground-Dove, Alopecoenas stairi
- Tanna Ground-Dove, Alopecoenas ferruginea
- Norfolk Ground-Dove, Alopecoenas norfolciensis
- Crested Pigeon, Ocyphaps lophotes
- Chestnut-quilled Rock-Pigeon, Petrophassa rufipennis
- White-quilled Rock-Pigeon, Petrophassa albipennis
- Wonga Pigeon, Leucosarcia melanoleuca
- Diamond Dove, Geopelia cuneata
- Zebra Dove, Geopelia striata
- Peaceful Dove, Geopelia placida
- Barred Dove, Geopelia maugeus
- Bar-shouldered Dove, Geopelia humeralis
- Common Bronzewing, Phaps chalcoptera
- Brush Bronzewing, Phaps elegans
- Flock Bronzewing, Phaps histrionica
- Spinifex Pigeon, Geophaps plumifera
- Squatter Pigeon, Geophaps scripta
- Partridge Pigeon, Geophaps smithii
Raphini Wetmore, 1930 (1835)
- Thick-billed Ground-Pigeon, Trugon terrestris
- Choiseul Pigeon, Microgoura meeki
- Pheasant Pigeon, Otidiphaps nobilis
- Western Crowned-Pigeon, Goura cristata
- Southern Crowned-Pigeon, Goura scheepmakeri
- Victoria Crowned-Pigeon, Goura victoria
- Tooth-billed Pigeon, Didunculus strigirostris
- Nicobar Pigeon, Caloenas nicobarica
- Spotted Green Pigeon, Caloenas maculata
- Dodo, Raphus cucullatus
- Rodrigues Solitaire, Pezophaps solitaria
Treronini: Green-Pigeons G.R. Gray, 1840
- Cinnamon-headed Green-Pigeon, Treron fulvicollis
- Little Green-Pigeon, Treron olax
- Pink-necked Green-Pigeon, Treron vernans
- Orange-breasted Green-Pigeon, Treron bicinctus
- Sri Lanka Green-Pigeon, Treron pompadora
- Gray-fronted Green-Pigeon, Treron affinis
- Ashy-headed Green-Pigeon, Treron phayrei
- Andaman Green-Pigeon, Treron chloropterus
- Philippine Green-Pigeon, Treron axillaris
- Buru Green-Pigeon, Treron aromaticus
- Thick-billed Green-Pigeon, Treron curvirostra
- Gray-cheeked Green-Pigeon, Treron griseicauda
- Sumba Green-Pigeon, Treron teysmannii
- Flores Green-Pigeon, Treron floris
- Timor Green-Pigeon, Treron psittaceus
- Large Green-Pigeon, Treron capellei
- Yellow-footed Green-Pigeon, Treron phoenicopterus
- Bruce's Green-Pigeon, Treron waalia
- Madagascan Green-Pigeon, Treron australis
- Comoros Green-Pigeon, Treron griveaudi
- African Green-Pigeon, Treron calvus
- Pemba Green-Pigeon, Treron pembaensis
- Sao Tome Green-Pigeon, Treron sanctithomae
- Pin-tailed Green-Pigeon, Treron apicauda
- Sumatran Green-Pigeon, Treron oxyurus
- Yellow-vented Green-Pigeon, Treron seimundi
- Wedge-tailed Green-Pigeon, Treron sphenurus
- White-bellied Green-Pigeon, Treron sieboldii
- Whistling Green-Pigeon, Treron formosae
Turturini G.R. Gray, 1840
- White-eared Brown-Dove, Phapitreron leucotis
- Amethyst Brown-Dove, Phapitreron amethystinus
- Tawitawi Brown-Dove, Phapitreron cinereiceps
- Mindanao Brown-Dove, Phapitreron brunneiceps
- Common Emerald-Dove, Chalcophaps indica
- Pacific Emerald-Dove, Chalcophaps longirostris
- Stephan's Emerald-Dove, Chalcophaps stephani
- Namaqua Dove, Oena capensis
- Emerald-spotted Wood-Dove, Turtur chalcospilos
- Black-billed Wood-Dove, Turtur abyssinicus
- Blue-spotted Wood-Dove, Turtur afer
- Tambourine Dove, Turtur tympanistria
- Blue-headed Wood-Dove, Turtur brehmeri
Ptilinopini: Fruit-Doves and Imperial-Pigeons Selby, 1835
- Pink-bellied Imperial-Pigeon, Ducula poliocephala
- White-bellied Imperial-Pigeon, Ducula forsteni
- Mindoro Imperial-Pigeon, Ducula mindorensis
- Gray-headed Imperial-Pigeon, Ducula radiata
- Spotted Imperial-Pigeon, Ducula carola
- Green Imperial-Pigeon, Ducula aenea
- Nicobar Imperial-Pigeon, Ducula nicobarica
- Spectacled Imperial-Pigeon, Ducula perspicillata
- Seram Imperial-Pigeon, Ducula neglecta
- Elegant Imperial-Pigeon, Ducula concinna
- Pacific Imperial-Pigeon, Ducula pacifica
- Micronesian Imperial-Pigeon, Ducula oceanica
- Polynesian Imperial-Pigeon, Ducula aurorae
- Marquesan Imperial-Pigeon, Ducula galeata
- Red-knobbed Imperial-Pigeon, Ducula rubricera
- Spice Imperial-Pigeon, Ducula myristicivora
- Purple-tailed Imperial-Pigeon, Ducula rufigaster
- Cinnamon-bellied Imperial-Pigeon, Ducula basilica
- Finsch's Imperial-Pigeon, Ducula finschii
- Rufescent Imperial-Pigeon, Ducula chalconota
- Island Imperial-Pigeon, Ducula pistrinaria
- Pink-headed Imperial-Pigeon, Ducula rosacea
- Christmas Imperial-Pigeon, Ducula whartoni
- Gray Imperial-Pigeon, Ducula pickeringii
- Barking Imperial-Pigeon, Ducula latrans
- Chestnut-bellied Imperial-Pigeon, Ducula brenchleyi
- Vanuatu Imperial-Pigeon, Ducula bakeri
- Goliath Imperial-Pigeon, Ducula goliath
- Pinon's Imperial-Pigeon, Ducula pinon
- Black Imperial-Pigeon, Ducula melanochroa
- Collared Imperial-Pigeon, Ducula mullerii
- Zoe's Imperial-Pigeon, Ducula zoeae
- Mountain Imperial-Pigeon, Ducula badia
- Dark-backed Imperial-Pigeon, Ducula lacernulata
- Timor Imperial-Pigeon, Ducula cineracea
- Pied Imperial-Pigeon, Ducula bicolor
- Silver-tipped Imperial-Pigeon, Ducula luctuosa
- Torresian Imperial-Pigeon, Ducula spilorrhoa
- Yellowish Imperial-Pigeon, Ducula subflavescens
- New Zealand Pigeon, Hemiphaga novaeseelandiae
- Chatham Pigeon, Hemiphaga chathamensis
- Topknot Pigeon, Lopholaimus antarcticus
- Sombre Pigeon, Cryptophaps poecilorrhoa
- Papuan Mountain-Pigeon, Gymnophaps albertisii
- Buru Mountain-Pigeon, Gymnophaps mada
- Seram Mountain-Pigeon, Gymnophaps stalkeri
- Pale Mountain-Pigeon, Gymnophaps solomonensis
- Scarlet-breasted Fruit-Dove, Ptilinopus bernsteinii
Click for Ptilinopus tree - Wompoo Fruit-Dove, Ptilinopus magnificus
- Flame-breasted Fruit-Dove, Ptilinopus marchei
- Cream-breasted Fruit-Dove, Ptilinopus merrilli
- Yellow-breasted Fruit-Dove, Ptilinopus occipitalis
- Red-eared Fruit-Dove, Ptilinopus fischeri
- Jambu Fruit-Dove, Ptilinopus jambu
- Oberholser's Fruit-Dove, Ptilinopus epius
- Banggai Fruit-Dove, Ptilinopus subgularis
- Sula Fruit-Dove, Ptilinopus mangoliensis
- Black-chinned Fruit-Dove, Ptilinopus leclancheri
- Black-naped Fruit-Dove, Ptilinopus melanospilus
- Golden Fruit-Dove, Ptilinopus luteovirens
- Orange Fruit-Dove, Ptilinopus victor
- Whistling Fruit-Dove, Ptilinopus layardi
- Mauritius Blue-Pigeon, Ptilinopus nitidissimus
- Madagascan Blue-Pigeon, Ptilinopus madagascariensis
- Comoros Blue-Pigeon, Ptilinopus sganzini
- Seychelles Blue-Pigeon, Ptilinopus pulcherrimus
- Pink-headed Fruit-Dove, Ptilinopus porphyreus
- Banded Fruit-Dove, Ptilinopus cinctus
- Red-naped Fruit-Dove, Ptilinopus dohertyi
- Black-banded Fruit-Dove, Ptilinopus alligator
- Dwarf Fruit-Dove, Ptilinopus nainus
- Cloven-feathered Dove, Ptilinopus holosericeus
- Superb Fruit-Dove, Ptilinopus superbus
- White-bibbed Fruit-Dove, Ptilinopus rivoli
- Yellow-bibbed Fruit-Dove, Ptilinopus solomonensis
- Tanna Fruit-Dove, Ptilinopus tannensis
- Orange-bellied Fruit-Dove, Ptilinopus iozonus
- Claret-breasted Fruit-Dove, Ptilinopus viridis
- White-headed Fruit-Dove, Ptilinopus eugeniae
- Knob-billed Fruit-Dove, Ptilinopus insolitus
- Gray-headed Fruit-Dove, Ptilinopus hyogastrus
- Carunculated Fruit-Dove, Ptilinopus granulifrons
- Pink-spotted Fruit-Dove, Ptilinopus perlatus
- Wallace's Fruit-Dove, Ptilinopus wallacii
- Orange-fronted Fruit-Dove, Ptilinopus aurantiifrons
- Ornate Fruit-Dove, Ptilinopus ornatus
- Beautiful Fruit-Dove, Ptilinopus pulchellus
- Blue-capped Fruit-Dove, Ptilinopus monacha
- Rose-crowned Fruit-Dove, Ptilinopus regina
- Coroneted Fruit-Dove, Ptilinopus coronulatus
- White-capped Fruit-Dove, Ptilinopus dupetithouarsii
- Red-moustached Fruit-Dove, Ptilinopus mercierii
- Red-bellied Fruit-Dove, Ptilinopus greyi
- Crimson-crowned Fruit-Dove, Ptilinopus porphyraceus
- Palau Fruit-Dove, Ptilinopus pelewensis
- Mariana Fruit-Dove, Ptilinopus roseicapilla
- Silver-capped Fruit-Dove, Ptilinopus richardsii
- Many-colored Fruit-Dove, Ptilinopus perousii
- Lilac-crowned Fruit-Dove, Ptilinopus rarotongensis
- Gray-green Fruit-Dove, Ptilinopus purpuratus
- Rapa Fruit-Dove, Ptilinopus huttoni
- Scarlet-capped Fruit-Dove, Ptilinopus insularis
- Makatea Fruit-Dove, Ptilinopus chalcurus
- Atoll Fruit-Dove, Ptilinopus coralensis
- Negros Fruit-Dove, Ptilinopus arcanus