Grallariidae: Antpittas Sclater and Salvin, 1873 (1872)
4 genera, 54 species Not HBW Family
The Urrao Antpitta, Grallaria urraoensis, has been named twice. The first formal description was by Barrera and Bartels (2010) using the name Grallaria fenwickorum. The SACC considers this description inadequate, (SACC #479B). They rely on the description by the original discoverers in Carantón and Certuche (2010), who use the name Grallaria urraoensis. There are allegations and controversy surrounding the unfortunate circumstances that led to the two different names. I won't further comment on that. Normally, the name Grallaria fenwickorum would have priority, being published first. However, since the SACC considers the proposed type speciment to be inadequate to identify the bird, they use the second published name, urraoensis. I follow the SACC decision in this matter.
It has been suspected that the Spotted Antpitta is more than one species (Ridgely and Tudor, 1994; Hilty, 2003; Krabbe and Schulenberg, 2003). Carneiro et al. (2012) made a close study of this. Based on their results, three species have been split from Spotted Antpitta, Hylopezus macularius:
- Zimmer's Antpitta, Hylopezus dilutus
- Alta Floresta Antpitta, Hylopezus whittakeri
- Snethlage's Antpitta, Hylopezus paraensis
The subspecies names differ a bit from Krabbe and Schulenberg (2003). Besides the newly created whittakeri, Carneiro et al. resurrect the name dilutus, previously considered a junior synonym of paraensis. They also submerged diversus into dilutus. Zimmer's Antpitta ranges north of the Amazon from the Rio Negro up into southern Venzuela and west into Colombia and Peru; the Alta Floresta Antpitta is restricted to the Madeira/Xingu area south of the Amazon; the Spotted Antpitta occurs on and around the Guianan shield; finally Snethlage's Antpitta occurs south of the Amazon and east of the Xingu.
- Ochre-breasted Antpitta, Grallaricula flavirostris
- Scallop-breasted Antpitta, Grallaricula loricata
- Hooded Antpitta, Grallaricula cucullata
- Peruvian Antpitta, Grallaricula peruviana
- Ochre-fronted Antpitta, Grallaricula ochraceifrons
- Rusty-breasted Antpitta, Grallaricula ferrugineipectus
- Slate-crowned Antpitta / Slaty-crowned Antpitta, Grallaricula nana
- Sucre Antpitta, Grallaricula cumanensis
- Crescent-faced Antpitta, Grallaricula lineifrons
- Thrush-like Antpitta, Myrmothera campanisona
- Tepui Antpitta, Myrmothera simplex
- Streak-chested Antpitta, Hylopezus perspicillatus
- Spotted Antpitta, Hylopezus macularius
- Zimmer's Antpitta, Hylopezus dilutus
- Alta Floresta Antpitta, Hylopezus whittakeri
- Snethlage's Antpitta, Hylopezus paraensis
- Masked Antpitta, Hylopezus auricularis
- Thicket Antpitta, Hylopezus dives
- White-lored Antpitta, Hylopezus fulviventris
- Amazonian Antpitta, Hylopezus berlepschi
- White-browed Antpitta, Hylopezus ochroleucus
- Speckle-breasted Antpitta, Hylopezus nattereri
- Undulated Antpitta, Grallaria squamigera
- Giant Antpitta, Grallaria gigantea
- Great Antpitta, Grallaria excelsa
- Variegated Antpitta, Grallaria varia
- Moustached Antpitta, Grallaria alleni
- Scaled Antpitta, Grallaria guatimalensis
- Tachira Antpitta, Grallaria chthonia
- Plain-backed Antpitta, Grallaria haplonota
- Ochre-striped Antpitta, Grallaria dignissima
- Elusive Antpitta, Grallaria eludens
- Chestnut-crowned Antpitta, Grallaria ruficapilla
- Watkins's Antpitta, Grallaria watkinsi
- Santa Marta Antpitta, Grallaria bangsi
- Cundinamarca Antpitta, Grallaria kaestneri
- Stripe-headed Antpitta, Grallaria andicolus
- Gray-naped Antpitta, Grallaria griseonucha
- Bicolored Antpitta, Grallaria rufocinerea
- Jocotoco Antpitta, Grallaria ridgelyi
- Chestnut-naped Antpitta, Grallaria nuchalis
- Pale-billed Antpitta, Grallaria carrikeri
- White-throated Antpitta, Grallaria albigula
- Yellow-breasted Antpitta, Grallaria flavotincta
- White-bellied Antpitta, Grallaria hypoleuca
- Rusty-tinged Antpitta, Grallaria przewalskii
- Bay Antpitta, Grallaria capitalis
- Red-and-white Antpitta, Grallaria erythroleuca
- Rufous Antpitta, Grallaria rufula
- Chestnut Antpitta, Grallaria blakei
- Tawny Antpitta, Grallaria quitensis
- Urrao Antpitta, Grallaria urraoensis
- Brown-banded Antpitta, Grallaria milleri
- Rufous-faced Antpitta, Grallaria erythrotis
Rhinocryptidae: Tapaculos Wetmore, 1926 (1837)
12 genera, 59 species HBW-8
The overall structure of the Rhinocryptidae is a combination of Maurício et al., 2008, Moyle et al. (2009), and Ericson et al. (2010). Only Ericson et al. examined enough taxa to accurately place Psilorhamphus. It is clear from both Ericson et al. and Moyle et al. that Liosceles through Teledromas form a clade (clade 1), as do Merulaxis through Scytalopus (clade 2). What is not clear is where the Scelorchilus/Pteroptochos goes. Moyle et al. place it sister to clade 2, while Ericson et al. prefer a tree with it sister to clade 1. In fact, when you look inside Ericson et al. to examine the individual gene trees, one places it in clade 1, one places it sister to clade 2, and the third puts it in a polytomy with clades 1 and 2. I don't think there is enough information to decide the issue, and have left it unresolved by putting Scelorchilus/Pteroptochos in a trichotomy with clades 1 and 2.
The arrangement of subfamilies, tribes, and subtribes used by Maurício et al. (2012) do not match the phylogeny used here (nor that of Moyle et al., 2009; nor Ericson et al., 2010). I consider that an artifact of their morphological analysis.
In version 2.04, the White-breasted and Bahia Tapaculos were moved out of Scytalopus into a new genus, Eleoscytalopus (see Maurício et al., 2008). The Diamantina Tapaculo was added in 2.05. The composition of Scytalopus is currently very contentious. The SACC discussion of the Diamantina Tapaculo demonstrates how complex the issues are.
Based on Maurício et al. (2014), the Boa Nova Tapaculo, Scytalopus gonzagai, has been split from the Mouse-colored Tapaculo, Scytalopus speluncae.
The Rock Tapaculo, Scytalopus petrophilus, was noted by Bornschein et al. (2007) and included in the genetic analysis of Mata et al. (2009). It was formally described by Whitney et al. (2010). The names of these taxa are in dispute, with some (Raposo et al., 2006, 2008) contending that speluncae does not apply to the Mouse-colored Tapaculo, but that it may apply to petrophilus. Raposo et al. (2006) introduced the alternate name notorius for the Mouse-colored Tapaculo. Maurício et al. (2010) argue in favor of the standard treatment, which is followed by the SACC and here.
To clarify a bit, regardless of the scientific names, the Mouse-colored Tapaculo (aka notorius) is the dark gray species (or species group) of more coastal mountains from Rio Grande do Sul to at least Minas Gerais and Espírito Santo, including the Serra do Caparaó, Serra do Mar, and Serra da Mantiqueira, while the Rock Tapaculo (aka petrophilus) is the light gray form in the interior range, Serra do Espinhaço.
The issue concerning the name is that the type specimen of speluncae was supposedly collected at São João del Rei, which is Rock Tapaculo territory. However, there are doubts whether this is correct, and others argue that in fact the specimen is a Mouse-colored Tapaculo with the actual collection location unknown. The specimen, collected by Ménétriés, is not in good shape, and access to it has been limited. The issues are whether Ménétriés was in error about the location for this specimen, and whether it is in fact a Mouse-colored Tapaculo. I'm following the SACC here, not because I'm entirely convinced they've made the right decision, but because to do otherwise would only futher complicate the issue.
The newly described Junin Tapaculo, Scytalopus gettyae, has been added (see Hosner et al., 2013). This bird has been known of for a while as the Satipo form of Large-footed Tapaculo and is not to be confused with the so-called Millpo Tapaculo, which occurs higher up. It appears to be part of the latrans group.
The newly described Perija Tapaculo, Scytalopus perijanus, has been added to the list (Avendaño et al., 2015).
- Rusty-belted Tapaculo, Liosceles thoracicus
- Spotted Bamboowren, Psilorhamphus guttatus
- Ocellated Tapaculo, Acropternis orthonyx
- Crested Gallito, Rhinocrypta lanceolata
- Sandy Gallito, Teledromas fuscus
- White-throated Tapaculo, Scelorchilus albicollis
- Chucao Tapaculo, Scelorchilus rubecula
- Chestnut-throated Huet-huet, Pteroptochos castaneus
- Black-throated Huet-huet, Pteroptochos tarnii
- Moustached Turca, Pteroptochos megapodius
- Slaty Bristlefront, Merulaxis ater
- Stresemann's Bristlefront, Merulaxis stresemanni
- White-breasted Tapaculo, Eleoscytalopus indigoticus
- Bahia Tapaculo, Eleoscytalopus psychopompus
- Ash-colored Tapaculo, Myornis senilis
- Ochre-flanked Tapaculo, Eugralla paradoxa
- Blackish Tapaculo, Scytalopus latrans
- Unicolored Tapaculo, Scytalopus unicolor
- Junin Tapaculo, Scytalopus gettyae
- Trilling Tapaculo, Scytalopus parvirostris
- Boa Nova Tapaculo, Scytalopus gonzagai
- Mouse-colored Tapaculo, Scytalopus speluncae
- Marsh Tapaculo, Scytalopus iraiensis
- Rock Tapaculo, Scytalopus petrophilus
- Brasilia Tapaculo, Scytalopus novacapitalis
- Diamantina Tapaculo, Scytalopus diamantinensis
- Planalto Tapaculo, Scytalopus pachecoi
- Large-footed Tapaculo, Scytalopus macropus
- Santa Marta Tapaculo, Scytalopus sanctaemartae
- Long-tailed Tapaculo, Scytalopus micropterus
- Rufous-vented Tapaculo, Scytalopus femoralis
- White-crowned Tapaculo / Northern White-crowned Tapaculo, Scytalopus atratus
- Bolivian Tapaculo / Bolivian White-crowned Tapaculo, Scytalopus bolivianus
- Tacarcuna Tapaculo, Scytalopus panamensis
- Choco Tapaculo, Scytalopus chocoensis
- Silvery-fronted Tapaculo, Scytalopus argentifrons
- Magdalena Tapaculo, Scytalopus rodriguezi
- Stiles's Tapaculo, Scytalopus stilesi
- Ecuadorian Tapaculo / El Oro Tapaculo, Scytalopus robbinsi
- Narino Tapaculo, Scytalopus vicinior
- Brown-rumped Tapaculo, Scytalopus latebricola
- Perija Tapaculo, Scytalopus perijanus
- Merida Tapaculo, Scytalopus meridanus
- Caracas Tapaculo, Scytalopus caracae
- Spillmann's Tapaculo, Scytalopus spillmanni
- Chusquea Tapaculo, Scytalopus parkeri
- Magellanic Tapaculo, Scytalopus magellanicus
- Pale-bellied Tapaculo, Scytalopus griseicollis
- Neblina Tapaculo, Scytalopus altirostris
- Ancash Tapaculo, Scytalopus affinis
- Tschudi's Tapaculo, Scytalopus acutirostris
- Vilcabamba Tapaculo, Scytalopus urubambae
- Puna Tapaculo, Scytalopus simonsi
- Zimmer's Tapaculo, Scytalopus zimmeri
- White-browed Tapaculo, Scytalopus superciliaris
- Dusky Tapaculo, Scytalopus fuscus
- Paramillo Tapaculo, Scytalopus canus
- Paramo Tapaculo, Scytalopus opacus
- Diademed Tapaculo, Scytalopus schulenbergi
Formicariidae: Antthrushes G.R. Gray, 1840 (1825)
2 genera, 11 species HBW-8
- Rufous-capped Antthrush, Formicarius colma
- Black-faced Antthrush, Formicarius analis
- Rufous-fronted Antthrush, Formicarius rufifrons
- Black-headed Antthrush, Formicarius nigricapillus
- Rufous-breasted Antthrush, Formicarius rufipectus
- Short-tailed Antthrush, Chamaeza campanisona
- Striated Antthrush, Chamaeza nobilis
- Such's Antthrush / Cryptic Antthrush, Chamaeza meruloides
- Schwartz's Antthrush, Chamaeza turdina
- Rufous-tailed Antthrush, Chamaeza ruficauda
- Barred Antthrush, Chamaeza mollissima
Furnariidae: Ovenbirds G.R. Gray, 1840
68 genera, 308 species HBW-8
 At the family level, the taxonomy agrees with SACC, folding the woodcreepers
into the Furnariidae as the subfamily Dendrocolaptinae. This was necessary
because the woodcreepers are nested within the Furnariidae, as shown on the
diagram to the right. If they were separated, the Furnariidae would no longer
be monophyletic. The SACC now divides the Furnariidae into three subfamilies:
Sclerurinae (leaftossers and miners), Furnariinae (ovenbirds), and Dendrocolaptinae
(woodcreepers). I arrange them as Sclerurinae, Dendrocolaptinae, Furnariinae
because Furnariinae is much larger than Dendrocolaptinae.
At the family level, the taxonomy agrees with SACC, folding the woodcreepers
into the Furnariidae as the subfamily Dendrocolaptinae. This was necessary
because the woodcreepers are nested within the Furnariidae, as shown on the
diagram to the right. If they were separated, the Furnariidae would no longer
be monophyletic. The SACC now divides the Furnariidae into three subfamilies:
Sclerurinae (leaftossers and miners), Furnariinae (ovenbirds), and Dendrocolaptinae
(woodcreepers). I arrange them as Sclerurinae, Dendrocolaptinae, Furnariinae
because Furnariinae is much larger than Dendrocolaptinae.
Xenops
However, as you can see on the diagram, I've included one more subfamily: Xenopinae. This group consists of the three Xenops species. The problem here is that is in unclear whether Xenops is closer to the woodcreepers or true ovenbirds. According to some genes Xenops is sister to Dendrocolaptinae, while others put it sister to Furnariinae (see Fjeldså et al., 2005, 2007; Irestedt et al., 2009b; Moyle et al., 2009b; Derryberry et al., 2011). Fjeldså et al. (2005) tell a nice story that makes Xenops somewhat analogous to a piculet. I have some sympathy for this argument as they sometimes remind me of piculets. Moreover, the nuclear RAG-1 and RAG-2 genes that Moyle et al. rely on may change too slowly to resolve all of the find details of the tree. On the other hand, the other genes used by Derryberry et al. did not change the RAG picture. Further, the cytochrome-b tree in Fjeldså et al. (2005) puts Xenops with the Furnariinae. However, Fjeldså et al. (2005, 2007) both use additional genes and end up with Xenops near the woodcreepers. Irestedt et al. (2009b) agree in their overall tree, but the is considerable disagreement among the individual gene trees. Given the conflicting results, I think the best course of action for now is to put Xenops in its own subfamily to indicate the uncertainty, but put it closer to Furnariinae than Dendrocolaptinae pending further information.
The other complication with Xenops is that the Rufous-tailed Xenops does not belong in Xenops (Moyle et al., 2009b; Irestedt et al., 2009b). Fortunately, there is an available genus name: Microxenops (Chapman 1914). Accordingly, the Rufous-tailed Xenops becomes Microxenops milleri and moves to Pygarrhichadini. Megaxenops was also formerly considered part of this group, but Remsen (2003) had already noted that it is not closely related to Xenops.
Leaftossers
The Tawny-throated Leaftosser, Sclerurus mexicanus has been split into Tawny-throated Leaftosser, Sclerurus mexicanus (mexicanus + pullus) and Dusky Leaftosser, Sclerurus obscurior, Andean Leaftosser, Sclerurus andinus, and Amazonian Leaftosser, Sclerurus macconnelli (peruvianus, macconnelli, and bahiae) based on d'Horta et al., (2013). See also SACC Proposal #603
Woodcreepers
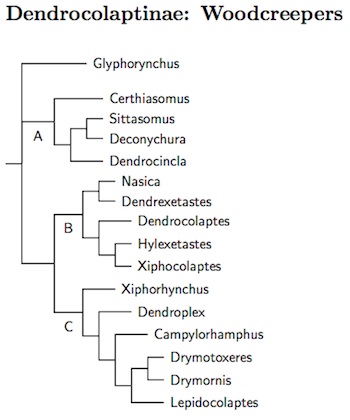 |
| Click for Dendrocolaptinae species |
|---|
We turn to the woodcreepers first. Irestedt et al. (2009b), Derryberry et al. (2011, 2012), and Moyle et al. (2009b) can be read as concurring that there are three clades (A, B, and C on the diagram), plus Glyphorynchus (Wedge-billed Woodcreeper). They agree on the membership and relative position of the three clades (A sister to B plus C). They are all consistent with the same genera, something only accomplished in the last few years. The big disagreement is the position of Glyphorynchus. Irestedt et al. and Derryberry et al. (2012) put it basal to the three clades, while Derryberry et al. (2011), Moyle et al. (2009b), and Ohlsen et al. (2013) have it sister to B plus C. What can one conclude? I gave up and put Glyphorynchus in an unresolved trichotomy with clade A and the combined clades B plus C. Glyphorynchus and A are sometimes characterized as “intermediate”, meaning between the other woodcreepers and the rest of Furnariidae, while B and C are sometimes referred to as “strong-billed” (e.g., HBW-8).
Within clade A, Deconychura has been split into two (Derryberry et al., 2010a). As a result the Spot-throated Woodcreeper is now Certhiasomus stictolaemus. It takes the basal position in clade A. I'm also treated Plain-winged Woodcreeper, Dendrocincla turdina as distinct from Plain-brown Woodcreeper, Dendrocincla fuliginosa (see Weir and Price, 2011; Derryberry et al., 2012). Note that taunayi then becomes a subspecies of Plain-winged Woodcreeper.
The strong-billed woodcreepers divide into a mostly heavy-billed group (B) and a clade contained the curved-billed species, including scythebills (C). In clade B, there's some disagreement about the placement of Dendrocolaptes. Derryberry et al. (2011, 2012) and Irestedt et al. (2009b) sample more taxa, and I follow them. Derryberry et al. (2012) also provide some support for Silva and Oren's (1995) treatment of Hylexetastes as four species. Accordingly, I've split Brigida's Woodcreeper, Hylexetastes brigidai, and Uniform Woodcreeper, Hylexetastes uniformis, from Red-billed Woodcreeper, Hylexetastes perrotii.
In the scythebill clade, the Greater Scythebill, formerly Campylorhamphus pucherani, turns out to be sister to the Scimitar-billed Woodcreeper, Drymornis bridgesii. Claramunt et al. (2010) established the new genus Drymotoxeres for it. Finally, there is some minor disagreement between Arbeláez-Cortés et al. (2012) and Derryberry et al. (2012) concerning the relationships of the basal three Lepidocolaptes. Derryberry et al. have them as closest relatives (as here), while the combined analysis of Arbeláez-Cortés et al. shows them as successive branches, in reversed order (the cytochrome-b agrees with Derryberry et al.).
Based on Rodrigues et al. (2013) and SACC Proposal #620, the Lineated Woodcreeper, Lepidocolaptes albolineatus, has been is split into 5 species. They are:
- White-lined Woodcreeper / Guianan Woodcreeper, Lepidocolaptes albolineatus
- Duida Woodcreeper, Lepidocolaptes duidae;
- Inambari Woodcreeper, Lepidocolaptes fatimalimae (new taxon);
- Rondonia Woodcreeper, Lepidocolaptes fuscicapillus (includes madeirae);
- Layard's Woodcreeper, Lepidocolaptes layardi.
True Ovenbirds
Once we get below the subfamily level, we abandon the early 2011 SACC order. The genera within the Furnariinae and Dendrocolaptinae are arranged quite differently than in the SACC list. The list here follows the recent tour-de-force by Derryberry and 8 co-authors (2011), which sampled all but 13 of the Furnariid species (96%). Before Derryberry et al., I had relied on a synthesis of Chesser et al. (2007), Fjeldså et al. (2005, 2007), Gonzalez and Wink (2008), Irestedt et al. (2004a, 2006a, 2009b), Moyle et al. (2009b), and Claramunt et al. (2010), with the overall arrangement is driven by the recent papers by Irestedt et al. (2009b) and Moyle et al. (2009b). I'm happy to say that I'm able to retain the same tribes (with the same members) as before Derryberry et al. The fact that this structure remains even though a number of new genes have been added to the analysis suggests that it is correct. There remain some minor issues concerning the arrangement of these tribes, both externally and internally, but the overall picture seems stable at this point.
As far as overall structure of the ovenbird subfamily is concerned, the main options are Irestedt et al. (2009b) topology found by Moyle et al. (2009b) and Derryberry et al. (2011). Derryberry et al. use six genes, two of which were previously used by Moyle et al. Irestedt et al. use an entirely different set of six genes and obtain a somewhat different topology. There Pygarrhichadini is the basal group, with Philydorini and Margarornini breaking off successively. A clade containing Berlepschiini, Furnariini, and Synallaxini is well-supported. A clade of Furnariini and Synallaxini get mediocre support, as does a division of Furnariini into two parts which are successively sister to Synallaxini. Except for the Pseudocolaptes, Premnornis, Tarphonomus group, there is agreement on the composition of the clades.
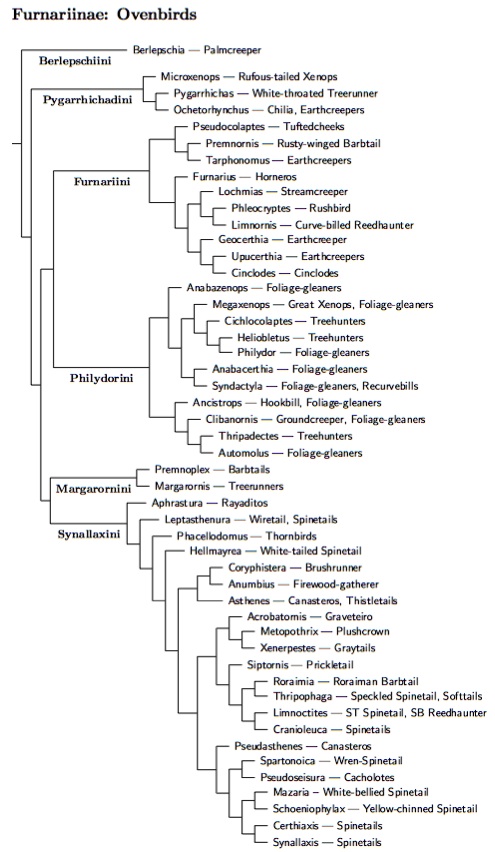 At the tribal level I now follow the topology of Moyle et al. (2009b) and Derryberry et al. (2011).
It starts with Berlepschia by itself at the base of the ovenbird
subfamily. I distinguish this and the major clades within the ovenbirds
by giving them tribal rank. You notice the various genera listed on the tree too. That
part of the tree, and the species trees below, are all based on Derryberry et al. (2011).
At the tribal level I now follow the topology of Moyle et al. (2009b) and Derryberry et al. (2011).
It starts with Berlepschia by itself at the base of the ovenbird
subfamily. I distinguish this and the major clades within the ovenbirds
by giving them tribal rank. You notice the various genera listed on the tree too. That
part of the tree, and the species trees below, are all based on Derryberry et al. (2011).
Berlepschiini is followed by another small tribe, Pygarrhichadini. Next comes the Furnariini-Philydorini group. It's followed by the Margarornini, which has been separated from the traditional Philydorinae. Finally, we have the Synallaxini.
There have also been some changes within these groups, sometimes involving reoganized genera. Note that the species composition of each tribe has remained the same since version 2.13 of this page (June 30, 2009). However, some have been moved to different genera.
The genus Upucerthia has been divided into 4 four parts (Chesser et al., 2007; Fjeldså et al. 2007). Two species are moved to Ochetorhynchus (in Pygarrhichadini), which also absorbs Chilia and Eremobius; two others form the new genus Tarphonomus (Chesser et al., 2007); U. serrana is placed in the new genus Geocerthia (Chesser et al., 2009); the rest remain in Upucerthia, which shrinks a bit with the lumping of Plain-breasted Earthcreeper, Upucerthia jelskii, into Buff-breasted Earthcreeper, Upucerthia validirostris (Areta and Pearman, 2013). The last three genera remain with the Furnariini.
This is not the only change in the Furnariini. The arrangement of Cinclodes species follows Derryberry et al. (2011), which is similar to that of Sanín et al. (2009). Sanín et al. presented evidence for splitting the Bar-winged Cinclodes, Cinclodes fuscus, into three species. Jaramillo (2003) suggested the English names Buff-winged Cinclodes for Cinclodes fuscus and Cream-winged Cinclodes for Cinclodes albiventris. SACC has named Cinclodes albidiventris Chestnut-winged Cinclodes. The Chestnut-winged Cinclodes is the northern group, with a range extending into NW Peru (Cajamarca and Piura). It includes subspecies heterurus and oreobates, in addition to albidiventris. The central species is the Cream-winged Cinclodes, ranging from N Peru (Amazonas) to N Chile (Antofagasta) and NW Argentina (La Rioja). It apparently includes the isolated subspecies riojanus, rufus, and yzurietaeof NW Argentina, as well as tucumanus and albiventris. The southern Buff-winged Cinclodes is then monotypic.
The Cipo Cinclodes, Cinclodes espinhacensis, is treated as a subspecies of the Long-tailed Cinclodes, Cinclodes pabsti. See SACC proposal #548.
There are several changes within Philydorini resulting from Derryberry et al. (2011). This includes the dismemberment of Philydor itself. Only 3 species remain in Philydor. The other 6 are reallocated as follows: The Ochre-breasted and Rufous-tailed Foliage-gleaners move to Anabacerthia, Buff-fronted and Chestnut-winged Foliage-gleaners move to Ancistrops, and Rufous-rumped and Slaty-winged Foliage-gleaners move to Megaxenops. The last two pairs of these are a bit unfortunate. The differences between the Chestnut-winged Hookbill and the two foliage-gleaners (Buff-fronted and Chestnut-winged) are fairly large, as are differences between the Great Xenops and Rufous-rumped and Slaty-winged Foliage-gleaners. However, I could not find available genus names belonging to any of the Buff-fronted, Chestnut-winged, Rufous-rumped, or Slaty-winged Foliage-gleaners.
Further, the two recurvebills Simoxenops have been moved into Syndactyla. They sit fairly deeply in Syndactyla, and even though they are distinctive, I don't see any other reasonable way to handle this.
The remaining changes to the Philydorini are confined to the Automolus branch. Chestnut-capped (Henna-capped) Foliage-gleaner is transferred from Hylocryptus to Clibanornis. The monotypic Hyloctistes has been merged into Automolus. Two Automolus species, the Ruddy and Santa Marta Foliage-gleaners, are placed in Clibanornis.
Finally, the newly discovered (and possibly extinct) Cryptic Treehunter, Cichlocolaptes mazarbarnetti, has been added. See Mazar Barnett and Buzzetti (2014) and Claramunt (2014b).
There were already changes in the Synallaxini prior to Derryberry et al. (2011). For example, Des Murs's Wiretail, formerly Sylviorthorhynchus desmursii, has been moved to the genus Leptasthenura.
The biggest changes in the Synallaxini involve the genera Asthenes, Oreophylax, and Schizoeaca. Here the usual generic limits do not reflect the gene tree. The recent paper by Derryberry et al. (2010b) has done much to straighten out the situation. Four of the Asthenes, humicola, patagonica, steinbachi, and cactorum, do not really belong to the group. Derryberry et al. (2010b) created the new genus Pseudasthenes for this clade. They are more closely related to Spartonoica and Pseudoseisura (Derryberry et al., 2010b; Fjeldså et al., 2007; Gonzalez and Wink, 2008; Irestedt et al., 2009b). This is a bit surprising as cactorum has been considered a subspecies of modesta, which is actually not a close relative.
The remaining species of the Asthenes, Oreophylax, and Schizoeaca group form a clade. Derryberry et al. (2010b) suggest treating them all as Asthenes, and I now follow that here.
There are 3 or 4 branches of Asthenes. The basal branch includes dorbignyi, baeri, and probably berlepschi. All of these have previously been considered part of a superspecies. Another supposed member of that group, luizae, appears to form a separate branch that is closer to the other Asthenes than to the dorbignyi superspecies. The dorbignyi group seems to include at least two undescribed taxa. One is the “Ancash” Canastero.
 |
 |
 |
| “Ancash” Canastero | ||
|---|---|---|
The remaining species of Asthenes are in two clades. The first runs from hudsoni to modesta. The other, which is comprised of the remaining Asthenes, includes the species formerly placed in Oreophylax and /Schizoeaca. Some of the relationships here are rather surprising as all of the old Schizoeaca are sometimes considered conspecific, and because of the geographic separation between the Andean Schizoeaca and Oreophylax of SE Brazil.
The Synallaxini have undergone further changes due to Derryberry et al. (2011). There's some reallocation in the Cranioleuca branch. Cranioleuca loses two species, Sulphur-throated Spinetail moves to Limnoctites and Speckled Spinetail moves to Thripophaga. Conversely, Thripophaga loses Russet-mantled Softtail to Cranioleuca. It also gains the newly discovered Delta Amacuro Softtail, Thripophaga amacurensis (Hilty et al., 2013).
I had previously separated several species of Synallaxis as Poecilurus in an attempt to avoid paraphyly. The more complete analysis of Derryberry et al. showed that this was a failure. According, I have returned the Poecilurus species to Synallaxis. Moreover, I've also merged the monotypic Gyalophylax and Siptornopsis into Synallaxis. Although Synallaxis is a large genus, the differences between the Synallaxis spinetails are small enough that I see no reason to subdivide it.
At the species level, the situation with the Cocoa (Xiphorhynchus susurrans), Buff-throated (X. guttatus), and Lafresnaye's (X. guttatoides) Woodcreepers is now clearer due to Rocha et al. (2015) and Aleixo (2001). I currently recognize 3 species in the complex.
- Lafresnaye's Woodcreeper, Xiphorhynchus guttatoides, including dorgibnyanus, eytoni, gracilirostris, and vicinalis (south and west of the Amazon/Negro).
- Cocoa Woodcreeper, Xiphorhynchus susurrans (Central America, Colombian Andes, northern Venezuela).
- Buff-throated Woodcreeper, Xiphorhynchus guttatus, including connectens and polystictus (north and east of the Amazon/Negro, Atlantic Forest).
The Cocoa and Buff-throated Woodcreepers are sister species, with the Lafresnaye's complex more distantly related. The Lafresnay'es complex may contain two additional species. However, as Rocha et al. (2015) point out, it would be prudent to sample upriver before accepting further splits. The dark-billed subspecies east of the Xingu (eytoni and gracilirostris) are sister taxa and only distantly related to the other subspecies of Lafresnaye's. They have sometimes been united as Dusky-billed Woodcreeper with the other dark-billed subspecies, vicinalis. However, the sampled vicinalis are quite a bit more closely related to guttatoides than to eytoni or gracilirostris. The subspecies vicinalis actually ranges from the Madeira to the Xingu (not to the Tapajós, as previously thought). It has been thought that some are intergrades between eytoni and guttatoides, but that was not supported by Rocha et al.'s samples.
Based on Derryberry et al. (2012) and Sousa-Neves et al. (2013), Tschudi's Woodcreeper, Xiphorhynchus chunchotambo, (including napensis and brevirostris) has been split from Ocellated Woodcreeper, Xiphorhynchus ocellatus. I follow the arrangement in Sousa-Neves et al., where the Chestnut-rumped Woodcreeper, Xiphorhynchus pardalotus, is basal to both.
The Bahia Spinetail, Synallaxis whitneyi, has been merged into the Rufous-capped Spinetail, Synallaxis ruficapilla as the characteristics of whitneyi are all within the variation in ruficapilla (Stopiglia et al., 2013).
Finally, the White-bellied Spinetail has been placed in the new genus Mazaria (Claramunt, 2014a) as Mazaria propinqua. SACC still lists it in Synallaxis, but genetic data (Derryberry et al., 2011) showed it was sister to Schoeniophylax and I moved it there. As Claramunt (2014a) notes, the two species are so different that putting the White-bellied Spinetail in Schoeniophylax makes the genus undiagnosible. He established the new genus Mazaria to solve this problem.
Sclerurinae: Leaftossers and Miners Swainson, 1827
- Short-billed Leaftosser, Sclerurus rufigularis
Click for Sclerurinae
species - Tawny-throated Leaftosser, Sclerurus mexicanus
- Dusky Leaftosser, Sclerurus obscurior
- Andean Leaftosser, Sclerurus andinus
- Amazonian Leaftosser, Sclerurus macconnelli
- Scaly-throated Leaftosser, Sclerurus guatemalensis
- Black-tailed Leaftosser, Sclerurus caudacutus
- Gray-throated Leaftosser, Sclerurus albigularis
- Rufous-breasted Leaftosser, Sclerurus scansor
- Coastal Miner, Geositta peruviana
- Slender-billed Miner, Geositta tenuirostris
- Common Miner, Geositta cunicularia
- Puna Miner, Geositta punensis
- Campo Miner, Geositta poeciloptera
- Thick-billed Miner, Geositta crassirostris
- Rufous-banded Miner, Geositta rufipennis
- Grayish Miner, Geositta maritima
- Short-billed Miner, Geositta antarctica
- Dark-winged Miner, Geositta saxicolina
- Creamy-rumped Miner, Geositta isabellina
Dendrocolaptinae: Woodcreepers G.R. Gray, 1840
- Wedge-billed Woodcreeper, Glyphorynchus spirurus
Click for Dendrocolaptinae
species - Spot-throated Woodcreeper, Certhiasomus stictolaemus
- Olivaceous Woodcreeper, Sittasomus griseicapillus
- Long-tailed Woodcreeper, Deconychura longicauda
- Tyrannine Woodcreeper, Dendrocincla tyrannina
- White-chinned Woodcreeper, Dendrocincla merula
- Ruddy Woodcreeper, Dendrocincla homochroa
- Tawny-winged Woodcreeper, Dendrocincla anabatina
- Plain-winged Woodcreeper, Dendrocincla turdina
- Plain-brown Woodcreeper, Dendrocincla fuliginosa
- Long-billed Woodcreeper, Nasica longirostris
- Cinnamon-throated Woodcreeper, Dendrexetastes rufigula
- Northern Barred-Woodcreeper, Dendrocolaptes sanctithomae
- Amazonian Barred-Woodcreeper, Dendrocolaptes certhia
- Planalto Woodcreeper, Dendrocolaptes platyrostris
- Black-banded Woodcreeper, Dendrocolaptes picumnus
- Hoffmanns's Woodcreeper, Dendrocolaptes hoffmannsi
- Brigida's Woodcreeper, Hylexetastes brigidai
- Uniform Woodcreeper, Hylexetastes uniformis
- Bar-bellied Woodcreeper, Hylexetastes stresemanni
- Red-billed Woodcreeper, Hylexetastes perrotii
- White-throated Woodcreeper, Xiphocolaptes albicollis
- Moustached Woodcreeper, Xiphocolaptes falcirostris
- Strong-billed Woodcreeper, Xiphocolaptes promeropirhynchus
- Great Rufous Woodcreeper, Xiphocolaptes major
- Lesser Woodcreeper, Xiphorhynchus fuscus
- Elegant Woodcreeper, Xiphorhynchus elegans
- Spix's Woodcreeper, Xiphorhynchus spixii
- Chestnut-rumped Woodcreeper, Xiphorhynchus pardalotus
- Tschudi's Woodcreeper, Xiphorhynchus chunchotambo
- Ocellated Woodcreeper, Xiphorhynchus ocellatus
- Spotted Woodcreeper, Xiphorhynchus erythropygius
- Olive-backed Woodcreeper, Xiphorhynchus triangularis
- Striped Woodcreeper, Xiphorhynchus obsoletus
- Ivory-billed Woodcreeper, Xiphorhynchus flavigaster
- Black-striped Woodcreeper, Xiphorhynchus lachrymosus
- Lafresnaye's Woodcreeper, Xiphorhynchus guttatoides
- Cocoa Woodcreeper, Xiphorhynchus susurrans
- Buff-throated Woodcreeper, Xiphorhynchus guttatus
- Straight-billed Woodcreeper, Dendroplex picus
- Zimmer's Woodcreeper, Dendroplex kienerii
- Black-billed Scythebill, Campylorhamphus falcularius
- Brown-billed Scythebill, Campylorhamphus pusillus
- Red-billed Scythebill, Campylorhamphus trochilirostris
- Curve-billed Scythebill, Campylorhamphus procurvoides
- Greater Scythebill, Drymotoxeres pucheranii
- Scimitar-billed Woodcreeper, Drymornis bridgesii
- Scaled Woodcreeper, Lepidocolaptes squamatus
- Scalloped Woodcreeper, Lepidocolaptes falcinellus
- Montane Woodcreeper, Lepidocolaptes lacrymiger
- White-striped Woodcreeper, Lepidocolaptes leucogaster
- Spot-crowned Woodcreeper, Lepidocolaptes affinis
- Streak-headed Woodcreeper, Lepidocolaptes souleyetii
- Narrow-billed Woodcreeper, Lepidocolaptes angustirostris
- White-lined Woodcreeper / Guianan Woodcreeper, Lepidocolaptes albolineatus
- Duida Woodcreeper, Lepidocolaptes duidae
- Inambari Woodcreeper, Lepidocolaptes fatimalimae
- Rondonia Woodcreeper, Lepidocolaptes fuscicapillus
- Layard's Woodcreeper, Lepidocolaptes layardi
Xenopinae: Xenops Bonaparte, 1854
- Plain Xenops, Xenops minutus
- Streaked Xenops, Xenops rutilans
- Slender-billed Xenops, Xenops tenuirostris
Furnariinae: True Ovenbirds G.R. Gray, 1840
Berlepschiini: Palmcreeper Ohlson et al., 2013a
- Point-tailed Palmcreeper, Berlepschia rikeri
Pygarrhichadini Wolters, 1977
- Rufous-tailed Xenops, Microxenops milleri
- White-throated Treerunner, Pygarrhichas albogularis
- Rock Earthcreeper, Ochetorhynchus andaecola
- Crag Chilia, Ochetorhynchus melanurus
- Straight-billed Earthcreeper, Ochetorhynchus ruficaudus
- Band-tailed Earthcreeper, Ochetorhynchus phoenicurus
Furnariini G.R. Gray, 1840
- Buffy Tuftedcheek, Pseudocolaptes lawrencii
Click for Xenopinae thru
Furnariini species - Streaked Tuftedcheek, Pseudocolaptes boissonneautii
- Rusty-winged Barbtail, Premnornis guttuliger
- Bolivian Earthcreeper, Tarphonomus harterti
- Chaco Earthcreeper, Tarphonomus certhioides
- Pale-billed Hornero / Bay Hornero, Furnarius torridus
- Pale-legged Hornero, Furnarius leucopus
- Wing-banded Hornero / Band-tailed Hornero, Furnarius figulus
- Lesser Hornero, Furnarius minor
- Rufous Hornero, Furnarius rufus
- Crested Hornero, Furnarius cristatus
- Sharp-tailed Streamcreeper, Lochmias nematura
- Wren-like Rushbird, Phleocryptes melanops
- Curve-billed Reedhaunter, Limnornis curvirostris
- Striated Earthcreeper, Geocerthia serrana
- Buff-breasted Earthcreeper, Upucerthia validirostris
- Patagonian Forest Earthcreeper, Upucerthia saturatior
- White-throated Earthcreeper, Upucerthia albigula
- Scale-throated Earthcreeper / Scaly-throated Earthcreeper, Upucerthia dumetaria
- Long-tailed Cinclodes, Cinclodes pabsti
- Buff-winged Cinclodes, Cinclodes fuscus
- Blackish Cinclodes, Cinclodes antarcticus
- Cordoba Cinclodes, Cinclodes comechingonus
- Chestnut-winged Cinclodes, Cinclodes albidiventris
- Olrog's Cinclodes, Cinclodes olrogi
- Cream-winged Cinclodes, Cinclodes albiventris
- Gray-flanked Cinclodes, Cinclodes oustaleti
- Stout-billed Cinclodes, Cinclodes excelsior
- Royal Cinclodes, Cinclodes aricomae
- White-bellied Cinclodes, Cinclodes palliatus
- White-winged Cinclodes, Cinclodes atacamensis
- Dark-bellied Cinclodes, Cinclodes patagonicus
- Surf Cinclodes / Peruvian Seaside Cinclodes, Cinclodes taczanowskii
- Seaside Cinclodes / Chilean Seaside Cinclodes, Cinclodes nigrofumosus
Philydorini: Foliage-gleaners, Treehunters Sclater & Salvin, 1873
- Dusky-cheeked Foliage-gleaner / Bamboo Foliage-gleaner, Anabazenops dorsalis
Click for Philydorini
and Margarornini species - White-collared Foliage-gleaner, Anabazenops fuscus
- Great Xenops, Megaxenops parnaguae
- Slaty-winged Foliage-gleaner, Megaxenops fuscipennis
- Rufous-rumped Foliage-gleaner, Megaxenops erythrocercus
- Cryptic Treehunter, Cichlocolaptes mazarbarnetti
- Pale-browed Treehunter, Cichlocolaptes leucophrus
- Sharp-billed Treehunter, Heliobletus contaminatus
- Cinnamon-rumped Foliage-gleaner, Philydor pyrrhodes
- Alagoas Foliage-gleaner, Philydor novaesi
- Black-capped Foliage-gleaner, Philydor atricapillus
- Montane Foliage-gleaner, Anabacerthia striaticollis
- Scaly-throated Foliage-gleaner, Anabacerthia variegaticeps
- Rufous-tailed Foliage-gleaner, Anabacerthia ruficaudata
- White-browed Foliage-gleaner, Anabacerthia amaurotis
- Ochre-breasted Foliage-gleaner, Anabacerthia lichtensteini
- Buff-browed Foliage-gleaner, Syndactyla rufosuperciliata
- Russet-mantled Foliage-gleaner / Planalto Foliage-gleaner, Syndactyla dimidiata
- White-throated Foliage-gleaner / Tepui Foliage-gleaner, Syndactyla roraimae
- Lineated Foliage-gleaner, Syndactyla subalaris
- Rufous-necked Foliage-gleaner, Syndactyla ruficollis
- Guttulate Foliage-gleaner, Syndactyla guttulata
- Peruvian Recurvebill, Syndactyla ucayalae
- Bolivian Recurvebill, Syndactyla striata
- Chestnut-winged Hookbill, Ancistrops strigilatus
- Buff-fronted Foliage-gleaner, Ancistrops rufus
- Chestnut-winged Foliage-gleaner, Ancistrops erythropterus
- Chestnut-capped Foliage-gleaner / Henna-capped Foliage-gleaner, Clibanornis rectirostris
- Canebrake Groundcreeper, Clibanornis dendrocolaptoides
- Santa Marta Foliage-gleaner, Clibanornis rufipectus
- Ruddy Foliage-gleaner, Clibanornis rubiginosus
- Henna-hooded Foliage-gleaner, Clibanornis erythrocephalus
- Uniform Treehunter, Thripadectes ignobilis
- Flammulated Treehunter, Thripadectes flammulatus
- Rufous-backed Treehunter / Peruvian Treehunter, Thripadectes scrutator
- Striped Treehunter, Thripadectes holostictus
- Streak-capped Treehunter, Thripadectes virgaticeps
- Streak-breasted Treehunter, Thripadectes rufobrunneus
- Black-billed Treehunter, Thripadectes melanorhynchus
- Chestnut-crowned Foliage-gleaner, Automolus rufipileatus
- Brown-rumped Foliage-gleaner, Automolus melanopezus
- Striped Woodhaunter / Eastern Woodhaunter, Automolus subulatus
- Buff-throated Foliage-gleaner, Automolus ochrolaemus
- Olive-backed Foliage-gleaner, Automolus infuscatus
- Para Foliage-gleaner, Automolus paraensis
- White-eyed Foliage-gleaner, Automolus leucophthalmus
- Pernambuco Foliage-gleaner, Automolus lammi
Margarornini: Barbtails and Treerunners Ridgway, 1911
- Spotted Barbtail, Premnoplex brunnescens
- White-throated Barbtail, Premnoplex tatei
- Ruddy Treerunner, Margarornis rubiginosus
- Pearled Treerunner, Margarornis squamiger
- Beautiful Treerunner, Margarornis bellulus
- Fulvous-dotted Treerunner / Star-chested Treerunner, Margarornis stellatus
Synallaxini: Spinetails de Selys-Longchamps, 1839 (1836)
- Masafuera Rayadito, Aphrastura masafuerae
Click for Synallaxini
species (4 pages) - Thorn-tailed Rayadito, Aphrastura spinicauda
- Des Murs's Wiretail, Leptasthenura desmurii
- Tawny Tit-Spinetail, Leptasthenura yanacensis
- White-browed Tit-Spinetail, Leptasthenura xenothorax
- Araucaria Tit-Spinetail, Leptasthenura setaria
- Tufted Tit-Spinetail, Leptasthenura platensis
- Striolated Tit-Spinetail, Leptasthenura striolata
- Rusty-crowned Tit-Spinetail, Leptasthenura pileata
- Streaked Tit-Spinetail / Streak-backed Tit-Spinetail, Leptasthenura striata
- Brown-capped Tit-Spinetail, Leptasthenura fuliginiceps
- Andean Tit-Spinetail, Leptasthenura andicola
- Plain-mantled Tit-Spinetail, Leptasthenura aegithaloides
- Rufous-fronted Thornbird, Phacellodomus rufifrons
- Streak-fronted Thornbird, Phacellodomus striaticeps
- Little Thornbird, Phacellodomus sibilatrix
- Chestnut-backed Thornbird, Phacellodomus dorsalis
- Spot-breasted Thornbird, Phacellodomus maculipectus
- Greater Thornbird, Phacellodomus ruber
- Freckle-breasted Thornbird, Phacellodomus striaticollis
- Orange-eyed Thornbird, Phacellodomus erythrophthalmus
- Orange-breasted Thornbird, Phacellodomus ferrugineigula
- White-browed Spinetail, Hellmayrea gularis
- Lark-like Brushrunner, Coryphistera alaudina
- Firewood-gatherer, Anumbius annumbi
- Short-billed Canastero, Asthenes baeri
- Berlepsch's Canastero, Asthenes berlepschi
- Creamy-breasted Canastero / Rusty-vented Canastero, Asthenes dorbignyi
- Cipo Canastero, Asthenes luizae
- Hudson's Canastero, Asthenes hudsoni
- Austral Canastero, Asthenes anthoides
- Line-fronted Canastero, Asthenes urubambensis
- Many-striped Canastero, Asthenes flammulata
- Junin Canastero, Asthenes virgata
- Scribble-tailed Canastero, Asthenes maculicauda
- Streak-backed Canastero, Asthenes wyatti
- Puna Canastero, Asthenes sclateri
- Streak-throated Canastero, Asthenes humilis
- Cordilleran Canastero, Asthenes modesta
- Itatiaia Spinetail, Asthenes moreirae
- Black-throated Thistletail, Asthenes harterti
- Sharp-billed Canastero, Asthenes pyrrholeuca
- Ochre-browed Thistletail, Asthenes coryi
- Perija Thistletail, Asthenes perijana
- White-chinned Thistletail, Asthenes fuliginosa
- Mouse-colored Thistletail, Asthenes griseomurina
- Puna Thistletail, Asthenes helleri
- Eye-ringed Thistletail, Asthenes palpebralis
- Rusty-fronted Canastero, Asthenes ottonis
- Canyon Canastero, Asthenes pudibunda
- Vilcabamba Thistletail, Asthenes vilcabambae
- Maquis Canastero, Asthenes heterura
- Pink-legged Graveteiro, Acrobatornis fonsecai
- Orange-fronted Plushcrown, Metopothrix aurantiaca
- Double-banded Graytail, Xenerpestes minlosi
- Equatorial Graytail, Xenerpestes singularis
- Spectacled Prickletail, Siptornis striaticollis
- Roraiman Barbtail, Roraimia adusta
- Speckled Spinetail, Thripophaga gutturata
- Plain Softtail, Thripophaga fusciceps
- Orinoco Softtail, Thripophaga cherriei
- Delta Amacuro Softtail, Thripophaga amacurensis
- Striated Softtail, Thripophaga macroura
- Straight-billed Reedhaunter, Limnoctites rectirostris
- Sulphur-throated Spinetail / Sulphur-bearded Spinetail, Limnoctites sulphuriferus
- Marcapata Spinetail, Cranioleuca marcapatae
- Light-crowned Spinetail, Cranioleuca albiceps
- Parker's Spinetail, Cranioleuca vulpecula
- Russet-mantled Softtail, Cranioleuca berlepschi
- Rusty-backed Spinetail, Cranioleuca vulpina
- Scaled Spinetail, Cranioleuca muelleri
- Creamy-crested Spinetail, Cranioleuca albicapilla
- Olive Spinetail, Cranioleuca obsoleta
- Pallid Spinetail, Cranioleuca pallida
- Stripe-crowned Spinetail, Cranioleuca pyrrhophia
- Bolivian Spinetail, Cranioleuca henricae
- Red-faced Spinetail, Cranioleuca erythrops
- Baron's Spinetail, Cranioleuca baroni
- Ash-browed Spinetail, Cranioleuca curtata
- Line-cheeked Spinetail, Cranioleuca antisiensis
- Coiba Spinetail, Cranioleuca dissita
- Tepui Spinetail, Cranioleuca demissa
- Gray-headed Spinetail, Cranioleuca semicinerea
- Streak-capped Spinetail, Cranioleuca hellmayri
- Crested Spinetail, Cranioleuca subcristata
- Dusky-tailed Canastero, Pseudasthenes humicola
- Patagonian Canastero, Pseudasthenes patagonica
- Steinbach's Canastero, Pseudasthenes steinbachi
- Cactus Canastero, Pseudasthenes cactorum
- Bay-capped Wren-Spinetail, Spartonoica maluroides
- Rufous Cacholote / Gray-crested Cacholote, Pseudoseisura unirufa
- Caatinga Cacholote, Pseudoseisura cristata
- Brown Cacholote, Pseudoseisura lophotes
- White-throated Cacholote, Pseudoseisura gutturalis
- White-bellied Spinetail, Mazaria propinqua
- Chotoy Spinetail, Schoeniophylax phryganophilus
- Yellow-chinned Spinetail, Certhiaxis cinnamomeus
- Red-and-white Spinetail, Certhiaxis mustelinus
- Ochre-cheeked Spinetail, Synallaxis scutata
- Gray-bellied Spinetail, Synallaxis cinerascens
- Plain-crowned Spinetail, Synallaxis gujanensis
- Maranon Spinetail, Synallaxis maranonica
- White-lored Spinetail, Synallaxis albilora
- Hoary-throated Spinetail, Synallaxis kollari
- Rufous-breasted Spinetail, Synallaxis erythrothorax
- White-whiskered Spinetail, Synallaxis candei
- Blackish-headed Spinetail, Synallaxis tithys
- Rusty-headed Spinetail, Synallaxis fuscorufa
- Rufous Spinetail, Synallaxis unirufa
- Black-throated Spinetail, Synallaxis castanea
- Stripe-breasted Spinetail, Synallaxis cinnamomea
- Ruddy Spinetail, Synallaxis rutilans
- Chestnut-throated Spinetail, Synallaxis cherriei
- Great Spinetail, Synallaxis hypochondriaca
- Necklaced Spinetail, Synallaxis stictothorax
- Russet-bellied Spinetail, Synallaxis zimmeri
- Slaty Spinetail, Synallaxis brachyura
- Silvery-throated Spinetail, Synallaxis subpudica
- Red-shouldered Spinetail, Synallaxis hellmayri
- Dusky Spinetail, Synallaxis moesta
- McConnell's Spinetail, Synallaxis macconnelli
- Cabanis's Spinetail, Synallaxis cabanisi
- Rufous-capped Spinetail, Synallaxis ruficapilla
- Pinto's Spinetail, Synallaxis infuscata
- Cinereous-breasted Spinetail, Synallaxis hypospodia
- Spix's Spinetail, Synallaxis spixi
- Dark-breasted Spinetail, Synallaxis albigularis
- Rio Orinoco Spinetail, Synallaxis beverlyae
- Pale-breasted Spinetail, Synallaxis albescens
- Sooty-fronted Spinetail, Synallaxis frontalis
- Azara's Spinetail, Synallaxis azarae
- Apurimac Spinetail, Synallaxis courseni