PICIFORMES Linnaeus, 1758
 |
| Click for Piciformes tree |
|---|
Linnaeus used order Picae and the genus Picus. Seems like a slam-dunk! Then why do others attribute Piciformes to Meyer and Wolf, 1810?
The Piciformes fossil record is shorter than that of the other orders in Afroaves. The Sylphornithidae are thought to be stem Piciformes. They are known from Europe starting in the middle Eocene. Recognizable Pici fossils are first found in the early Oligocene.
GALBULI Vigors, 1825
The jacamars and puffbirds are more closely related to each other than to the rest of the Piciformes. They are sometimes placed in their own order, Galbuliformes. The higher-level taxonomy of the Piciformes is based on Ericson et al. (2006a), Johansson and Ericson (2003) and Moyle (2004). Details of the Galbulidae and Bucconidae are from Witt (2004), who included most of the species in his analysis.
Galbulidae: Jacamars Vigors, 1825
5 genera, 19 species HBW-7
The Spot-tailed Jacamar, Galbula rufoviridis (including heterogyna), has been split from the Rufous-tailed Jacamar, Galbula ruficauda. There are indications that further splits might be needed. See Witt (2004).
- Three-toed Jacamar, Jacamaralcyon tridactyla
Click for Galbulidae tree - Dusky-backed Jacamar, Brachygalba salmoni
- Pale-headed Jacamar, Brachygalba goeringi
- Brown Jacamar, Brachygalba lugubris
- White-throated Jacamar, Brachygalba albogularis
- Great Jacamar, Jacamerops aureus
- White-eared Jacamar, Galbalcyrhynchus leucotis
- Purus Jacamar, Galbalcyrhynchus purusianus
- Paradise Jacamar, Galbula dea
- Purplish Jacamar, Galbula chalcothorax
- Bronzy Jacamar, Galbula leucogastra
- Yellow-billed Jacamar, Galbula albirostris
- Blue-cheeked Jacamar / Blue-necked Jacamar, Galbula cyanicollis
- Rufous-tailed Jacamar, Galbula ruficauda
- Coppery-chested Jacamar, Galbula pastazae
- Green-tailed Jacamar, Galbula galbula
- White-chinned Jacamar, Galbula tombacea
- Bluish-fronted Jacamar, Galbula cyanescens
- Spot-tailed Jacamar, Galbula rufoviridis
Bucconidae: Puffbirds Horsfield, 1821
The Chestnut-capped Puffbird is placed in genus Cyphos (Spix 1824) rather than Argicus (Cabanis and Heine 1863) because Cyphos is not preoccupied by Cyphus under current ICZN rules.
12 genera, 35 species HBW-7
- Rusty-breasted Nunlet, Nonnula rubecula
Click for Bucconidae tree - Rufous-capped Nunlet, Nonnula ruficapilla
- Chestnut-headed Nunlet, Nonnula amaurocephala
- Gray-cheeked Nunlet, Nonnula frontalis
- Brown Nunlet, Nonnula brunnea
- Fulvous-chinned Nunlet, Nonnula sclateri
- Black-streaked Puffbird, Malacoptila fulvogularis
- White-whiskered Puffbird, Malacoptila panamensis
- Moustached Puffbird, Malacoptila mystacalis
- Crescent-chested Puffbird, Malacoptila striata
- Rufous-necked Puffbird, Malacoptila rufa
- White-chested Puffbird, Malacoptila fusca
- Semicollared Puffbird, Malacoptila semicincta
- Collared Puffbird, Bucco capensis
- Spot-backed Puffbird / Caatinga Puffbird, Nystalus maculatus
- Barred Puffbird, Nystalus radiatus
- Striolated Puffbird, Nystalus striolatus
- White-eared Puffbird, Nystalus chacuru
- Swallow-winged Puffbird / Swallow-wing, Chelidoptera tenebrosa
- Yellow-billed Nunbird, Monasa flavirostris
- White-fronted Nunbird, Monasa morphoeus
- Black-fronted Nunbird, Monasa nigrifrons
- Black Nunbird, Monasa atra
- White-faced Nunbird, Hapaloptila castanea
- Lanceolated Monklet, Micromonacha lanceolata
- Chestnut-capped Puffbird, Cyphos macrodactylus
- Russet-throated Puffbird, Hypnelus ruficollis
- Sooty-capped Puffbird, Nystactes noanamae
- Spotted Puffbird, Nystactes tamatia
- Pied Puffbird, Notharchus tectus
- Brown-banded Puffbird, Notharchus ordii
- Black-breasted Puffbird, Notharchus pectoralis
- White-necked Puffbird, Notharchus hyperrhynchus
- Guianan Puffbird, Notharchus macrorhynchos
- Buff-bellied Puffbird, Notharchus swainsoni
PICI Linnaeus, 1758
Megalaimidae: Asian Barbets Blyth, 1852
2 genera, 34 species HBW-7 (split)
Barbet and Toucan taxonomy has not been completely resolved (see Barker and Lanyon, 2000; Moyle, 2004). The five families used here represent one possibility. Another is to roll all five into one family, Ramphastidae, as in the Howard and Moore checklist (Dickinson and Remsen, 2013). Another is to keep four of the five familes and merge the toucan-barbets into either the toucans (most likely) or New World barbets (possibly). The position of both Caloramphus and Trachyphonus among the other barbets has also not been completely resolved.
|
| Click for Megalaimidae tree |
|---|
Moyle (2004) found that the Fire-tufted Barbet is embedded deep within the traditional Megalaima. I have put them all in the same genus as a result. The genus name Psilopogon (Muller 1835, type pyrolophus) replaces Megalaima (G.R. Gray 1842, type virens) due to priority. Although this changes the type genus, the family name remains Megalaimidae.
More recently, den Tex and Leonard (2013) analyzed all of the Megalaimidae. The current species tree is based on their analysis. They recommended elevating several taxa to species level. Four of those recommendations are followed here. (1) The Sooty Barbet, Caloramphus hayii has been split from Brown Barbet, Caloramphus fuliginosus. (2) Yellow-eared Barbet, Psilopogon duvaucelii has been split from Blue-eared Barbet, Psilopogon australis. The later is considered monotypic. Since cyanotis and duvaucelii are thought to hybridize in Thailand, the remaining races are assigned to species duvaucelii. (3) The Golden-faced Barbet, Psilopogon chrysopsis (monotypic), has been split from Golden-whiskered Barbet, Psilopogon chrysopogon. (4) Finally, the Turquoise-throated Barbet, Psilopogon chersonesus, which is locally endenmic on the Kra Isthmus, has been split from the Blue-throated Barbet, Psilopogon asiatica. The names "Sooty Barbet", "Horsfield's Barbet", "Golden-faced Barbet", and "Kra Barbet" are used in the absence of any established English names.
Den Tex and Leonard (2013) also recommend splitting Psilopogon auricularis from Psilopogon franklinii. However, these taxa are believed to hybridize in Vietnam (Annam) and nearby Laos. Given that, I think additional information is needed to split them. They also note that Psilopogon asiatica likely contains at least one more species, but that further study is required to sort out the situation.
- Brown Barbet, Caloramphus fuliginosus
- Sooty Barbet, Caloramphus hayii
- Coppersmith Barbet, Psilopogon haemacephalus
- Malabar Barbet, Psilopogon malabaricus
- Crimson-fronted Barbet, Psilopogon rubricapillus
- Bornean Barbet, Psilopogon eximius
- Yellow-eared Barbet, Psilopogon duvaucelii
- Blue-eared Barbet, Psilopogon australis
- Fire-tufted Barbet, Psilopogon pyrolophus
- Great Barbet, Psilopogon virens
- Red-vented Barbet, Psilopogon lagrandieri
- Red-crowned Barbet, Psilopogon rafflesii
- Red-throated Barbet, Psilopogon mystacophanos
- Black-banded Barbet, Psilopogon javensis
- Golden-naped Barbet, Psilopogon pulcherrimus
- Yellow-crowned Barbet, Psilopogon henricii
- Flame-fronted Barbet, Psilopogon armillaris
- Green-eared Barbet, Psilopogon faiostrictus
- Lineated Barbet, Psilopogon lineatus
- Brown-headed Barbet, Psilopogon zeylanicus
- White-cheeked Barbet, Psilopogon viridis
- Yellow-fronted Barbet, Psilopogon flavifrons
- Golden-throated Barbet, Psilopogon franklinii
- Mountain Barbet, Psilopogon monticola
- Brown-throated Barbet, Psilopogon corvinus
- Golden-whiskered Barbet, Psilopogon chrysopogon
- Golden-faced Barbet, Psilopogon chrysopsis
- Moustached Barbet, Psilopogon incognitus
- Chinese Barbet, Psilopogon faber
- Taiwan Barbet, Psilopogon nuchalis
- Turquoise-throated Barbet, Psilopogon chersonesus
- Black-browed Barbet, Psilopogon oorti
- Indochinese Barbet, Psilopogon annamensis
- Blue-throated Barbet, Psilopogon asiaticus
Lybiidae: African Barbets, Tinkerbirds Sibley & Ahlquist, 1985
The 4th edition of the Howard and Moore checklist (Dickinson and Remsen, 2013) separated Pogonornis (Billberg 1828,type dubius) from Lybius. However, although they are somewhat sparse, the results in Moyle (2004) suggest that more changes are needed. The genus Tricholaema is restricted to the Hairy-breasted Barbet. Two former Tricholaema move to Lybius (Spot-flanked and Black-throated Barbets). The other three former Tricholaema form the genus Notopogonius (Roberts 1922, type leucomelas).
11 genera, 42 species HBW-7 (split)
- Yellow-billed Barbet, Trachylaemus purpuratus
- Crested Barbet, Trachyphonus vaillantii
- D'Arnaud's Barbet, Trachyphonus darnaudii
- Red-and-yellow Barbet, Trachyphonus erythrocephalus
- Yellow-breasted Barbet, Trachyphonus margaritatus
- Green Barbet, Cryptolybia olivacea
- Yellow-spotted Barbet, Buccanodon duchaillui
- Gray-throated Barbet, Gymnobucco bonapartei
- Sladen's Barbet, Gymnobucco sladeni
- Bristle-nosed Barbet, Gymnobucco peli
- Naked-faced Barbet, Gymnobucco calvus
- White-eared Barbet, Stactolaema leucotis
- Whyte's Barbet, Stactolaema whytii
- Anchieta's Barbet, Stactolaema anchietae
- Speckled Tinkerbird, Pogoniulus scolopaceus
- Green Tinkerbird, Pogoniulus simplex
- Moustached Tinkerbird, Pogoniulus leucomystax
- Western Tinkerbird, Pogoniulus coryphaea
- Red-rumped Tinkerbird, Pogoniulus atroflavus
- Yellow-throated Tinkerbird, Pogoniulus subsulphureus
- Yellow-rumped Tinkerbird, Pogoniulus bilineatus
- White-chested Tinkerbird, Pogoniulus makawai
- Red-fronted Tinkerbird, Pogoniulus pusillus
- Yellow-fronted Tinkerbird, Pogoniulus chrysoconus
- Hairy-breasted Barbet, Tricholaema hirsuta
- Red-fronted Barbet, Notopogonius diadematus
- Miombo Pied-Barbet, Notopogonius frontatus
- Acacia Pied-Barbet, Notopogonius leucomelas
- Brown-breasted Barbet, Pogonornis melanopterus
- Black-backed Barbet, Pogonornis minor
- Double-toothed Barbet, Pogonornis bidentatus
- Bearded Barbet, Pogonornis dubius
- Black-breasted Barbet, Pogonornis rolleti
- Spot-flanked Barbet, Lybius lacrymosus
- Black-throated Barbet, Lybius melanocephalus
- Banded Barbet, Lybius undatus
- Vieillot's Barbet, Lybius vieilloti
- White-headed Barbet, Lybius leucocephalus
- Chaplin's Barbet, Lybius chaplini
- Red-faced Barbet, Lybius rubrifacies
- Black-billed Barbet, Lybius guifsobalito
- Black-collared Barbet, Lybius torquatus
Capitonidae: New World Barbets Bonaparte, 1838
2 genera, 15 species HBW-7 (split)
The arrangement of species within Capito is based on the maximum likelihood tree in Armenta et al. (2005), which did not include the White-mantled Barbet or the recently discovered Sira Barbet (Seeholzer et al., 2012). Interestingly, this is the second recently discovered barbet in Peru, the other being the closely related Scarlet-banded Barbet (O'Neill et al., 2000). Although the genetic distance between them is fairly small, they have developed distinctive plumages are here treated as separate species.
- Lemon-throated Barbet, Eubucco richardsoni
- Scarlet-hooded Barbet, Eubucco tucinkae
- Red-headed Barbet, Eubucco bourcierii
- Versicolored Barbet, Eubucco versicolor
- Scarlet-crowned Barbet, Capito aurovirens
- Five-colored Barbet, Capito quinticolor
- Black-girdled Barbet, Capito dayi
- White-mantled Barbet, Capito hypoleucus
- Scarlet-banded Barbet, Capito wallacei
- Sira Barbet, Capito fitzpatricki
- Spot-crowned Barbet, Capito maculicoronatus
- Orange-fronted Barbet, Capito squamatus
- Black-spotted Barbet, Capito niger
- Gilded Barbet, Capito auratus
- Brown-chested Barbet, Capito brunneipectus
Semnornithidae: Toucan-barbets Prum, 1988
1 genus, 2 species HBW-7 (split)
- Prong-billed Barbet, Semnornis frantzii
- Toucan Barbet, Semnornis ramphastinus
Ramphastidae: Toucans, Aracaris, Toucanets Vigors, 1825
6 genera, 44 species HBW-7
 |
| Click for Ramphastidae Tree |
|---|
Moyle (2004) laid out the overall structure of the toucans used here, as shown in the tree diagram (Barker and Lanyon, 2000, is a bit different).
The age of the Ramphastidae crown-group is rather uncertain. Patel et al. (2011) estimate it at about 11 million years ago (late Miocene), while Lutz et al. (2013) estimate it about about 21 million years (early Miocene). Lutz et al. estimate the common ancestor with Semnornis at 29 mya (±5) while Patel et al. put it at 13 mya (±3).
The ordering of the Ramphastos toucans is based on Patané et al. (2009). The Toco Toucan is basal, and the rest fall into two sister groups, the smooth-billed yelping toucans (ambiguus through tucanus) and the channel-billed croaking toucans (sulfuratus through dicolorus). Although previous studies by Weckstein (2004, 2005) suggested the species limits needed adjustment, the analysis by Patané et al. (2009), which used more genes and taxa, did not concur.
The English name of Ramphastos ambiguus is changed to Black-mandibled Toucan / Yellow-throated Toucan. The point is that Black-mandibled properly applies only to the ambiguus group, but is not appropriate when swainsonii (Chestnut-mandibled Toucan) is included in the species. See SACC #663, and note that the NACC still uses Black-mandibled.
The aracari (Pteroglossus) sequence is based on Patel et al. (2011), which builds on Kimura et al. (2004) and Pereira and Wajntal (2008). The aracaris fall into several clades: (1) bailloni, viridis, and inscriptus (including humboldti); (2) torquatus (including nuchalis and erythrozonus), frantzii, sanguineus, and erythropygius; (3) azara (including flavirostris and mariae), bitorquatus and beauharnaesii; and (4) aracari, castanotis, and pluricinctus. Note that Pteroglossus bailloni, which has sometimes been considered a separate genus (Baillonius) is firmly embedded in Pteroglossus.
The subspecies P. azara mariae is sometimes separated as Brown-mandibled Aracari, leaving azara and flavirostris joined as Ivory-billed Aracari. However, Patel et al. (2011) found that mariae is more closely related to flavirostris than either is to azara. Since the genes don't match the usual grouping of these taxa, and since it is unclear whether they are separate biological species, I'm leaving them all lumped into Ivory-billed Aracari for now, although it is possible that 2 or 3 species are involved here.
I do not follow SACC concerning the Pteroglossus torquatus complex. Haffer (1967) found a narrow hybridization zone between P. torquatus and P. sanguineus (10-20 km across). This sort of hybridization zone is usually taken as evidence of biologically separate species. There is some weak support from Patel et al. (2011) who found no evidence of hybridization. However, their sample size is small and this cannot be taken as strong evidence. The genetic differences do not compel either way. Ridgely and Greenfield (2001) report limited hybridization between P. sanguineus and P. erythropygius near Playa de Oro, Ecuador, and Short and Horne (2001) report more extensive hybridization near Gualea. The exact size of the hybridization zone is not known, but it appears to be fairly limited. The result of all this is that I recognize Stripe-billed Aracari, Pteroglossus sanguineus, and Pale-mandibled Aracari, Pteroglossus erythropygius, as distinct species, pending more definitive data.
The sequence of the Andigena and Selenidera toucans is based on Lutz et al. (2013). The Yellow-eared Toucanet appears to group with the mountain-toucans rather than the other yellow-eared toucanets. Since no genus name is available, and Lutz et al. declined to create one, I'm listing it as "Selenidera" spectabilis. One alternative would be to include the mountain-toucans in Selenidera, another would be to move the Yellow-eared Toucanet into Andigena. Given distinctive appearance of the mountain-toucans, I think both are bad ideas. There is some question about whether the Guianan Toucanet belongs with the other yellow-eared toucanets (this is why Lutz et al. were reluctant to create a new genus).
Two molecular studies of the Aulacorhynchus toucanets have focused on different sets of species. Puebla-Olivares et al. (2008) concentrated on the emerald toucanet complex, while Bonaccorso et al. (2011) focus on the others.
Bonaccorso et al. (2011) found that the Chestnut-tipped Toucanet (Aulacorhynchus derbianus) contains two groups that are not each other's closest relatives. Following their recommendataions, it has been split into Whitely's Toucanet, Aulacorhynchus whitelianus (subspecies duidae, whitelianus, and osgoodi) and Derby's Toucanet, Aulacorhynchus derbianus (subspecies nigrirostris and derbianus). They also examined samples of all three subspecies of the Groove-billed Toucanet, Aulacorhynchus sulcatus. Although A. s. calorhynchus is sometimes considered a separate species (Yellow-billed Toucanet), it was nested within the sulcatus clade. The samples of calorhynchus formed a monophyletic group, suggesting that speciation is underway, but further study will be needed to clarify whether is has yet reached the point where it should be considered a separate biological species.
Some authors had previously considered the Emerald Toucanet to include multiple species (e.g., Navarro et al., 2001). The paper by Puebla-Olivares et al. (2008) considerably strengthens the case, and I have split them accordingly. Except for the treatment of albivitta and griseigularis as separate species, Clements 6th edition also follows the split. SACC has not yet considered the issue.
The Middle American portion of the Emerald Toucanet complex is fairly clearcut. This includes A. cognatus through A. prasinus with maxillaris included in A. caeruleogularis and all of the other Middle American subspecis in A. prasinus. The only real question is whether to lump A. wagleri into A. prasinus.
The South American Emerald Toucanets are another matter. There are some key gaps in the taxa analyzed by Puebla-Olivares et al., and the situation concerning albivitta is rather confusing. The races lautus (believed to be a distinct species) and phaeolaemus (probably closest to griseigularis) were not analyzed. Moreover, the samples from NE Ecuador, believed to be albivitta, do not appear to be albivitta. This creates a hole at the boundary between albivitta and atrogularis. For the present, atrogularis includes cyanolaemus and dimidiatus as subspecies.
Conventional wisdom has been that griseigularis and phaeolaemus group with albivitta. But Puebla-Olivares et al. found griseigularis in a tight grouping with atrogularis and the Ecuadorian ‘albivitta’. What is one to think? It could be that there is an un-named white-throated subspecies in Ecuador and perhaps southern Colombia. It could equally be that there was a glitch somewhere in the collection or analysis of the Ecuadorian ‘albivitta’. In any event, it makes the situation unclear. I have left A. griseigularis as a separate species pending a resolution of the problem, although I expect it to be folded into either A. atrogularis or A. albivitta.
- Toco Toucan, Ramphastos toco
- White-throated Toucan, Ramphastos tucanus
- Black-mandibled Toucan/Yellow-throated Toucan, Ramphastos ambiguus
- Keel-billed Toucan, Ramphastos sulfuratus
- Red-breasted Toucan / Green-billed Toucan, Ramphastos dicolorus
- Choco Toucan, Ramphastos brevis
- Channel-billed Toucan, Ramphastos vitellinus
- Saffron Toucanet, Pteroglossus bailloni
- Green Aracari, Pteroglossus viridis
- Lettered Aracari, Pteroglossus inscriptus
- Collared Aracari, Pteroglossus torquatus
- Fiery-billed Aracari, Pteroglossus frantzii
- Stripe-billed Aracari, Pteroglossus sanguineus
- Pale-mandibled Aracari, Pteroglossus erythropygius
- Ivory-billed Aracari, Pteroglossus azara
- Red-necked Aracari, Pteroglossus bitorquatus
- Curl-crested Aracari, Pteroglossus beauharnaesii
- Black-necked Aracari, Pteroglossus aracari
- Chestnut-eared Aracari, Pteroglossus castanotis
- Many-banded Aracari, Pteroglossus pluricinctus
- Yellow-eared Toucanet, "Selenidera" spectabilis
- Plate-billed Mountain-Toucan, Andigena laminirostris
- Black-billed Mountain-Toucan, Andigena nigrirostris
- Gray-breasted Mountain-Toucan, Andigena hypoglauca
- Hooded Mountain-Toucan, Andigena cucullata
- Guianan Toucanet, Selenidera piperivora
- Tawny-tufted Toucanet, Selenidera nattereri
- Golden-collared Toucanet, Selenidera reinwardtii
- Gould's Toucanet, Selenidera gouldii
- Spot-billed Toucanet, Selenidera maculirostris
- Yellow-browed Toucanet, Aulacorhynchus huallagae
- Blue-banded Toucanet, Aulacorhynchus coeruleicinctis
- Crimson-rumped Toucanet, Aulacorhynchus haematopygus
- Whitely's Toucanet, Aulacorhynchus whitelianus
- Groove-billed Toucanet, Aulacorhynchus sulcatus
- Derby's Toucanet, Aulacorhynchus derbianus
- Violet-throated Toucanet, Aulacorhynchus cognatus
- Blue-throated Toucanet, Aulacorhynchus caeruleogularis
- Emerald Toucanet, Aulacorhynchus prasinus
- Wagler's Toucanet, Aulacorhynchus wagleri
- Gray-throated Toucanet, Aulacorhynchus griseigularis
- Black-throated Toucanet, Aulacorhynchus atrogularis
- Santa Marta Toucanet, Aulacorhynchus lautus
- White-throated Toucanet, Aulacorhynchus albivitta
Indicatoridae: Honeyguides Swainson, 1837
4 genera, 17 species HBW-7
- Cassin's Honeybird, Prodotiscus insignis
- Green-backed Honeybird, Prodotiscus zambesiae
- Brown-backed Honeybird, Prodotiscus regulus
- Zenker's Honeyguide, Melignomon zenkeri
- Yellow-footed Honeyguide, Melignomon eisentrauti
- Dwarf Honeyguide, Indicator pumilio
- Willcocks's Honeyguide, Indicator willcocksi
- Pallid Honeyguide, Indicator meliphilus
- Least Honeyguide, Indicator exilis
- Thick-billed Honeyguide, Indicator conirostris
- Lesser Honeyguide, Indicator minor
- Spotted Honeyguide, Indicator maculatus
- Scaly-throated Honeyguide, Indicator variegatus
- Yellow-rumped Honeyguide, Indicator xanthonotus
- Malaysian Honeyguide, Indicator archipelagicus
- Greater Honeyguide, Indicator indicator
- Lyre-tailed Honeyguide, Melichneutes robustus
Picidae: Woodpeckers Leach, 1820
34 genera, 232 species HBW-7
 |
| Click for Picidae genera |
|---|
The details of woodpecker taxonomy are still being worked out, with species being shuffled around between genera (e.g., in Picoides and Veniliornis). The papers by Benz et al. (2006), Fuchs et al. (2006c, 2007b, 2008a), Fuchs and Pons (2015), Moore et al. (2006), Overton and Rhoads (2006), Webb and Moore (2005), and Weibel and Moore (2002a, 2002b) have all proven helpful.
This version is based on a closer look at the above and the summary of the Melanerpini by Raty. A certain amount of uncertainty remains, but I think this is a pretty good representation of the current state of woodpecker taxonomy. The diagram sums it up.
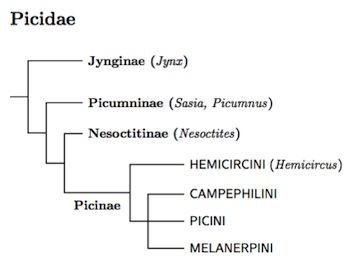
Woodpecker Phylogeny
That the wrynecks were sister to the remaining woodpeckers has been known for a while, as has the fact that the piculets were sister to the rest. Benz et al. (2006) showed that the Antillean Piculet does not belong with the other piculets, but is the sister of the Picinae. Accordingly, it is placed in the monotypic subfamily Nesoctitinae.
Fuchs et al. (2007b, 2008a) found that Hemicircus is sister to the rest of the Picinae. The TiF list now uses Fuchs et al. (2013) for the structure of the rest of the Picinae, but the relationship between the Campephilini, Picini, and Melanerpini remains unresolved.
 The key point that Fuchs et al. (2013) resolves is the relation between
Campephilus and an Asian clade containing the flamebacks. Morphology
has always suggested they are closely related, but genetic
analyses were yielding conflicting results. Fuchs et al. provide evidence that
this is due to a hybridization event that occured when the melanerpine
and Campephilus woodpeckers colonized the Americas.
They found that Blythipicus through Chrysocolaptes
is sister to Campephilus. We recognize the 4 genera together
as the tribe Campephilini.
The key point that Fuchs et al. (2013) resolves is the relation between
Campephilus and an Asian clade containing the flamebacks. Morphology
has always suggested they are closely related, but genetic
analyses were yielding conflicting results. Fuchs et al. provide evidence that
this is due to a hybridization event that occured when the melanerpine
and Campephilus woodpeckers colonized the Americas.
They found that Blythipicus through Chrysocolaptes
is sister to Campephilus. We recognize the 4 genera together
as the tribe Campephilini.
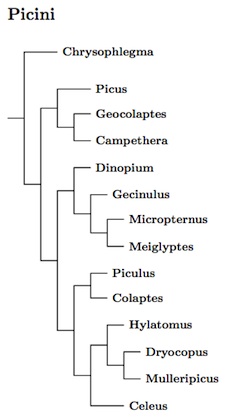
Picini seems to look something like the tree to the right (Fuchs et al., 2013). Here Celeus brachyurus has been placed in the monotypic genus Micropternus, Chrysophlegma has been separated from Picus, and Piculus rubiginosus and P. rivolii move to Colaptes. The order of species in Picus is based on a combination of Fuchs et al. (2008a) and Pons et al. (2011) while the ordering of Colaptes is based on Moore et al. (2011).
Contrary to studies using fewer genes, Dryocopus appears to be paraphyletic. It seems likely that the division is between New World and Old World species, with the Old World Dryocopus being closer to Mulleripicus. The New World Dryocopus then take the name Hylatomus (Baird 1858, type pileatus).
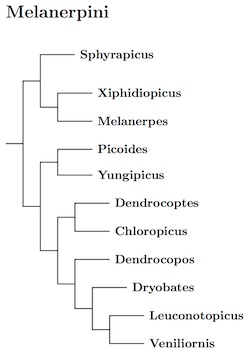 The genera Melanerpes, Sphyrapicus, and Xiphidiopicus
group together (see Overton and Rhoads, 2006 for details). That clade is
sister to the remaining Melanerpini. Previously, various papers have to be pieced
together to make a complete picture of the rest of Melanerpini. More recently, Fuchs
and Pons (2015) included enough taxa to form a complete picture. At the genus level
it turns out to be very similar to that I had previously pieced together, although
a couple of taxa had to be moved, and two genus names changed.
The genera Melanerpes, Sphyrapicus, and Xiphidiopicus
group together (see Overton and Rhoads, 2006 for details). That clade is
sister to the remaining Melanerpini. Previously, various papers have to be pieced
together to make a complete picture of the rest of Melanerpini. More recently, Fuchs
and Pons (2015) included enough taxa to form a complete picture. At the genus level
it turns out to be very similar to that I had previously pieced together, although
a couple of taxa had to be moved, and two genus names changed.
The genus Picoides has been split into three pieces: Picoides, Dryobates, and Leuconotopicus. It is not alone. The genus Dendrocopos has also been split into Dendrocopos, Yungipicus, and Dendrocoptes, but gains Hypopicus and Sapheopipo which are now subsumed in Dendrocopos (see Winkler et al., 2005). The remaining Picoides are sister to Yungipicus (formerly part of Dendrocopos), while the other two parts of the old Picoides (Leuconotopicus and Dryobates) are successive sisters to Veniliornis. The other piece of the former Dendrocopos, Dryobates, is sister to Dendropicos.
H&M-4 (Dickinson and Remsen, 2013) brought attention to the name Chloropicus (Malherbe 1845, type pyrrhogaster), which has priority over Dendropicos (Malherbe 1849, type fuscescens, subspecies lafresnayi), so the name Dendropicos is replaced by Chloropicus. With these changes, the TiF take on Melanerpini is shown in the diagram.
Further Issues
H&M-4 have a different approach to Chloropicus, treating it as four genera: Chloropicus, Dendropicos, Mesopicos, and Ipophilus. Although the taxon sampling is rather sparse, Fuchs and Pons (2015) find that Ipophilus is embeded in Mesopicos and suggests both taxa are embeded in Dendropicos. A further complication is that the Yellow-crowned Woodpecker, Leiopicus mahrattensis, is also in this group. The tree using the most data suggests Leiopicus (Bonaparte 1854, type mahrattensis) and Chloropicus are sisters, and that the three form a clade. However, there is also some support in the paper for putting Leiopicus as basal among the three with Chloropicus and Dendropicos sister. The ongoing study of this clade by Fuchs, Bowie, Carre and Pons should resolve the question. Given the current uncertainties, I prefer to lump them for under the name Chloropicus.
A few species besides those mentioned above have changed genera. The Arabian Woodpecker, Chloropicus dorae (formerly Dendropicos), has been moved into genus Dendrocoptes. Note that the name Dendrocoptes (Cabanis and Heine 1863, type medius), has priority over Desertipicus (Kinnear and Bates 1935, type dorae). Also, the Lesser Spotted Woodpecker, formerly Dendrocopos minor, is now in Dryobates; the Smoky-brown Woodpecker, Veniliornis fumigatus moved to Leuconotopicus; the Checkered and Striped Woodpeckers (formerly Picoides mixtus and P. lignarius) moved to Veniliornis.
Splits
Campephilini: The former Greater Flameback has been split into 6 species (Collar, 2011). Three of the new species are monotypic: haematribon, xanthocephalus, and erythrocephalus; while strictus includes kangeanensis and both rufopunctatus and montanus are included in lucidus. The remaining subspecies belong to guttacristatus, which retains the name Greater Flameback.
Picini: Following Collar (2011), the Philippine race of Common Flameback, Dinopium javanense, has been elevated to species status as Spot-throated Flameback, Dinopium everetti.
The Iberian Green Woodpecker, Picus sharpei, has been split from the European Green-Woodpecker, Picus viridis, based on Perktas et al. (2011) and Pons et al. (2011). The TiF list tries to use the biological species concept when possible. As pointed out by Perktas et al., the case for biological species status for the Zagros Green-Woodpecker remains weak, so it remains a subspecies, Picus viridis innominatus.
Moore et al. (2011) provided evidence that the Bronze-winged Woodpecker, Colaptes aeruginosus, of NE Mexico is sister to the Gray-crowned Woodpecker, Colaptes auricularis, rather than being a subspecies of the Golden-olive Woodpecker, Colaptes rubiginosus (which itself is sister to Black-necked Woodpecker, Colaptes atricollis).
As recommedned by Benz and Robbins (2011), I've split Ochre-backed Woodpecker, Celeus ochraceus, from Blond-crested Woodpecker, Celeus flavescens.
Melanerpini: A recent paper by García-Trejo et al. (2009) finds that the northern subspecies of the Golden-fronted Woodpecker, Melanerpes aurifrons, is more closely related to the Red-bellied Woodpecker, Melanerpes carolinus, than to other Golden-fronted races. Accordingly, they recommend splitting the other races as the tropical Melanerpes santacruzi, known as Velasquez's Woodpecker. It is possible that further splitting will be needed. The names Lesson's Woodpecker and Truxillo Woodpecker have been applied to some of the other tropical races. Based on their work, I have also rearranged the carolinus group, which starts at M. pucherani. The results of Overton and Rhoads (2006) suggest that the Puerto Rican, Hispaniolan, and presumably Jamaican Woodpeckers belong near carolinus. The entire Red-bellied complex needs further study to before it is completely sorted out.
Jynginae: Wrynecks Swainson, 1831
- Eurasian Wryneck, Jynx torquilla
- Red-throated Wryneck, Jynx ruficollis
Picumninae: Piculets G.R. Gray, 1840
- African Piculet, Sasia africana
- Rufous Piculet, Sasia abnormis
- White-browed Piculet, Sasia ochracea
- Speckled Piculet, Picumnus innominatus
- Bar-breasted Piculet, Picumnus aurifrons
- Orinoco Piculet, Picumnus pumilus
- Lafresnaye's Piculet, Picumnus lafresnayi
- Golden-spangled Piculet, Picumnus exilis
- Ecuadorian Piculet, Picumnus sclateri
- Scaled Piculet, Picumnus squamulatus
- White-bellied Piculet, Picumnus spilogaster
- Arrowhead Piculet, Picumnus minutissimus
- Spotted Piculet, Picumnus pygmaeus
- Speckle-chested Piculet, Picumnus steindachneri
- Varzea Piculet, Picumnus varzeae
- White-barred Piculet, Picumnus cirratus
- Ocellated Piculet, Picumnus dorbignyanus
- Ochre-collared Piculet, Picumnus temminckii
- White-wedged Piculet, Picumnus albosquamatus
- Rusty-necked Piculet, Picumnus fuscus
- Rufous-breasted Piculet, Picumnus rufiventris
- Tawny Piculet, Picumnus fulvescens
- Ochraceous Piculet, Picumnus limae
- Mottled Piculet, Picumnus nebulosus
- Plain-breasted Piculet, Picumnus castelnau
- Fine-barred Piculet, Picumnus subtilis
- Olivaceous Piculet, Picumnus olivaceus
- Grayish Piculet, Picumnus granadensis
- Chestnut Piculet, Picumnus cinnamomeus
Nesoctitinae: Antillean Piculet Wolters, 1976
- Antillean Piculet, Nesoctites micromegas
Picinae: Woodpeckers Leach, 1820
Hemicircini Cabanis and Heine, 1863
- Gray-and-buff Woodpecker, Hemicircus concretus
- Heart-spotted Woodpecker, Hemicircus canente
Campephilini Blyth, 1852
- Maroon Woodpecker, Blythipicus rubiginosus
- Bay Woodpecker, Blythipicus pyrrhotis
- Orange-backed Woodpecker, Reinwardtipicus validus
- Buff-spotted Flameback, Chrysocolaptes lucidus
- Luzon Flameback, Chrysocolaptes haematribon
- Yellow-faced Flameback, Chrysocolaptes xanthocephalus
- Red-headed Flameback, Chrysocolaptes erythrocephalus
- Javan Flameback, Chrysocolaptes strictus
- Greater Flameback, Chrysocolaptes guttacristatus
- Crimson-backed Flameback, Chrysocolaptes stricklandi
- White-naped Flameback / White-naped Woodpecker, Chrysocolaptes festivus
- Powerful Woodpecker, Campephilus pollens
- Crimson-bellied Woodpecker, Campephilus haematogaster
- Red-necked Woodpecker, Campephilus rubricollis
- Robust Woodpecker, Campephilus robustus
- Crimson-crested Woodpecker, Campephilus melanoleucos
- Pale-billed Woodpecker, Campephilus guatemalensis
- Guayaquil Woodpecker, Campephilus gayaquilensis
- Cream-backed Woodpecker, Campephilus leucopogon
- Magellanic Woodpecker, Campephilus magellanicus
- Ivory-billed Woodpecker, Campephilus principalis
- Imperial Woodpecker, Campephilus imperialis
Picini Leach, 1820
- Banded Woodpecker, Chrysophlegma miniaceum
- Checker-throated Woodpecker, Chrysophlegma mentale
- Greater Yellownape, Chrysophlegma flavinucha
- Lesser Yellownape, Picus chlorolophus
- Crimson-winged Woodpecker, Picus puniceus
- Japanese Green Woodpecker, Picus awokera
- Black-headed Woodpecker, Picus erythropygius
- Gray-headed Woodpecker, Picus canus
- Levaillant's Woodpecker, Picus vaillantii
- European Green-Woodpecker, Picus viridis
- Iberian Green-Woodpecker, Picus sharpei
- Scaly-bellied Woodpecker, Picus squamatus
- Streak-throated Woodpecker, Picus xanthopygaeus
- Red-collared Woodpecker, Picus rabieri
- Streak-breasted Woodpecker, Picus viridanus
- Laced Woodpecker, Picus vittatus
- Ground Woodpecker, Geocolaptes olivaceus
- Fine-spotted Woodpecker, Campethera punctuligera
- Bennett's Woodpecker, Campethera bennettii
- Speckle-throated Woodpecker, Campethera scriptoricauda
- Nubian Woodpecker, Campethera nubica
- Golden-tailed Woodpecker, Campethera abingoni
- Mombasa Woodpecker, Campethera mombassica
- Knysna Woodpecker, Campethera notata
- Green-backed Woodpecker, Campethera cailliautii
- Little Green Woodpecker, Campethera maculosa
- Tullberg's Woodpecker, Campethera tullbergi
- Buff-spotted Woodpecker, Campethera nivosa
- Brown-eared Woodpecker, Campethera caroli
- Olive-backed Woodpecker, Dinopium rafflesii
- Himalayan Flameback, Dinopium shorii
- Common Flameback, Dinopium javanense
- Spot-throated Flameback, Dinopium everetti
- Black-rumped Flameback, Dinopium benghalense
- Pale-headed Woodpecker, Gecinulus grantia
- Bamboo Woodpecker, Gecinulus viridis
- Rufous Woodpecker, Micropternus brachyurus
- Buff-rumped Woodpecker, Meiglyptes tristis
- Black-and-buff Woodpecker, Meiglyptes jugularis
- Buff-necked Woodpecker, Meiglyptes tukki
- Rufous-winged Woodpecker, Piculus simplex
- Stripe-cheeked Woodpecker, Piculus callopterus
- White-throated Woodpecker, Piculus leucolaemus
- Lita Woodpecker, Piculus litae
- Yellow-throated Woodpecker, Piculus flavigula
- Golden-green Woodpecker, Piculus chrysochloros
- White-browed Woodpecker / Yellow-browed Woodpecker, Piculus aurulentus
- Fernandina's Flicker, Colaptes fernandinae
- Northern Flicker, Colaptes auratus
- Gilded Flicker, Colaptes chrysoides
- Crimson-mantled Woodpecker, Colaptes rivolii
- Green-barred Woodpecker, Colaptes melanochloros
- Chilean Flicker, Colaptes pitius
- Andean Flicker, Colaptes rupicola
- Spot-breasted Woodpecker, Colaptes punctigula
- Campo Flicker, Colaptes campestris
- Gray-crowned Woodpecker, Colaptes auricularis
- Bronze-winged Woodpecker, Colaptes aeruginosus
- Golden-olive Woodpecker, Colaptes rubiginosus
- Black-necked Woodpecker, Colaptes atricollis
- Helmeted Woodpecker, Hylatomus galeatus
- Black-bodied Woodpecker, Hylatomus schulzii
- Lineated Woodpecker, Hylatomus lineatus
- Pileated Woodpecker, Hylatomus pileatus
- White-bellied Woodpecker, Dryocopus javensis
- Andaman Woodpecker, Dryocopus hodgei
- Black Woodpecker, Dryocopus martius
- Ashy Woodpecker, Mulleripicus fulvus
- Sooty Woodpecker, Mulleripicus funebris
- Great Slaty Woodpecker, Mulleripicus pulverulentus
- Cinnamon Woodpecker, Celeus loricatus
- Ringed Woodpecker, Celeus torquatus
- Ochre-backed Woodpecker, Celeus ochraceus
- Blond-crested Woodpecker, Celeus flavescens
- Chestnut Woodpecker, Celeus elegans
- Pale-crested Woodpecker, Celeus lugubris
- Chestnut-colored Woodpecker, Celeus castaneus
- Waved Woodpecker, Celeus undatus
- Scale-breasted Woodpecker / Scaly-breasted Woodpecker, Celeus grammicus
- Cream-colored Woodpecker, Celeus flavus
- Rufous-headed Woodpecker, Celeus spectabilis
- Kaempfer's Woodpecker, Celeus obrieni
Melanerpini G.R. Gray, 1846
- Williamson's Sapsucker, Sphyrapicus thyroideus
- Yellow-bellied Sapsucker, Sphyrapicus varius
- Red-naped Sapsucker, Sphyrapicus nuchalis
- Red-breasted Sapsucker, Sphyrapicus ruber
- Cuban Green Woodpecker, Xiphidiopicus percussus
- White Woodpecker, Melanerpes candidus
- Lewis's Woodpecker, Melanerpes lewis
- Guadeloupe Woodpecker, Melanerpes herminieri
- Red-headed Woodpecker, Melanerpes erythrocephalus
- Acorn Woodpecker, Melanerpes formicivorus
- Yellow-tufted Woodpecker, Melanerpes cruentatus
- Yellow-fronted Woodpecker, Melanerpes flavifrons
- Golden-naped Woodpecker, Melanerpes chrysauchen
- Beautiful Woodpecker, Melanerpes pulcher
- White-fronted Woodpecker, Melanerpes cactorum
- Gray-breasted Woodpecker, Melanerpes hypopolius
- Black-cheeked Woodpecker, Melanerpes pucherani
- Yucatan Woodpecker, Melanerpes pygmaeus
- Red-crowned Woodpecker, Melanerpes rubricapillus
- Hoffmann's Woodpecker, Melanerpes hoffmannii
- Golden-cheeked Woodpecker, Melanerpes chrysogenys
- Gila Woodpecker, Melanerpes uropygialis
- West Indian Woodpecker, Melanerpes superciliaris
- Puerto Rican Woodpecker, Melanerpes portoricensis
- Hispaniolan Woodpecker, Melanerpes striatus
- Jamaican Woodpecker, Melanerpes radiolatus
- Golden-fronted Woodpecker, Melanerpes aurifrons
- Velasquez's Woodpecker, Melanerpes santacruzi
- Red-bellied Woodpecker, Melanerpes carolinus
- Black-backed Woodpecker, Picoides arcticus
- Eurasian Three-toed Woodpecker, Picoides tridactylus
- American Three-toed Woodpecker, Picoides dorsalis
- Gray-capped Pygmy-Woodpecker, Yungipicus canicapillus
- Japanese Pygmy-Woodpecker, Yungipicus kizuki
- Sulawesi Pygmy-Woodpecker, Yungipicus temminckii
- Brown-capped Pygmy-Woodpecker, Yungipicus nanus
- Philippine Pygmy-Woodpecker, Yungipicus maculatus
- Sunda Pygmy-Woodpecker, Yungipicus moluccensis
- Sulu Pygmy-Woodpecker, Yungipicus ramsayi
- Middle Spotted Woodpecker, Dendrocoptes medius
- Arabian Woodpecker, Dendrocoptes dorae
- Brown-fronted Woodpecker, Dendrocoptes auriceps
- Yellow-crowned Woodpecker, Chloropicus mahrattensis
- Bearded Woodpecker, Chloropicus namaquus
- Yellow-crested Woodpecker, Chloropicus xantholophus
- Fire-bellied Woodpecker, Chloropicus pyrrhogaster
- Little Gray Woodpecker, Chloropicus elachus
- Speckle-breasted Woodpecker, Chloropicus poecilolaemus
- Abyssinian Woodpecker, Chloropicus abyssinicus
- Cardinal Woodpecker, Chloropicus fuscescens
- Gabon Woodpecker, Chloropicus gabonensis
- Melancholy Woodpecker, Chloropicus lugubris
- Stierling's Woodpecker, Chloropicus stierlingi
- Elliot's Woodpecker, Chloropicus elliotii
- African Gray-Woodpecker, Chloropicus goertae
- Eastern Gray-Woodpecker, Chloropicus spodocephalus
- Olive Woodpecker, Chloropicus griseocephalus
- Brown-backed Woodpecker, Chloropicus obsoletus
- Fulvous-breasted Woodpecker, Dendrocopos macei
- Freckle-breasted Woodpecker, Dendrocopos analis
- Stripe-breasted Woodpecker, Dendrocopos atratus
- Rufous-bellied Woodpecker, Dendrocopos hyperythrus
- White-backed Woodpecker, Dendrocopos leucotos
- Okinawa Woodpecker, Dendrocopos noguchii
- Darjeeling Woodpecker, Dendrocopos darjellensis
- White-winged Woodpecker, Dendrocopos leucopterus
- Great Spotted Woodpecker, Dendrocopos major
- Himalayan Woodpecker, Dendrocopos himalayensis
- Syrian Woodpecker, Dendrocopos syriacus
- Sind Woodpecker, Dendrocopos assimilis
- Lesser Spotted Woodpecker, Dryobates minor
- Crimson-breasted Woodpecker, Dryobates cathpharius
- Downy Woodpecker, Dryobates pubescens
- Ladder-backed Woodpecker, Dryobates scalaris
- Nuttall's Woodpecker, Dryobates nuttallii
- Red-cockaded Woodpecker, Leuconotopicus borealis
- Smoky-brown Woodpecker, Leuconotopicus fumigatus
- White-headed Woodpecker, Leuconotopicus albolarvatus
- Hairy Woodpecker, Leuconotopicus villosus
- Arizona Woodpecker, Leuconotopicus arizonae
- Strickland's Woodpecker, Leuconotopicus stricklandi
- White-spotted Woodpecker, Veniliornis spilogaster
- Checkered Woodpecker, Veniliornis mixtus
- Striped Woodpecker, Veniliornis lignarius
- Choco Woodpecker, Veniliornis chocoensis
- Golden-collared Woodpecker, Veniliornis cassini
- Red-rumped Woodpecker, Veniliornis kirkii
- Blood-colored Woodpecker, Veniliornis sanguineus
- Little Woodpecker, Veniliornis passerinus
- Dot-fronted Woodpecker, Veniliornis frontalis
- Scarlet-backed Woodpecker, Veniliornis callonotus
- Yellow-vented Woodpecker, Veniliornis dignus
- Bar-bellied Woodpecker, Veniliornis nigriceps
- Red-stained Woodpecker, Veniliornis affinis
- Yellow-eared Woodpecker, Veniliornis maculifrons