TELLURAVES Yuri et al., 2013
Telluraves contains slightly more than 3/4 of all bird species. Athough there's some residual uncertainty about the other main branches of Passerea, the land bird clade Telluraves is well-supported by Jarvis et al. (2014) where it receives 100% bootstrap support. Further, Hackett et al. (2008) and Ericson et al. (2006a) both found the hawks and American vultures to be closer to the Picimorphae than to the falcons (see also Suh et al., 2011; McCormack et al., 2013; Yuri et al. 2013). Other papers (e.g., Morgan-Richards et al., 2008) are less supportive, putting all of them somewhere between the Ardeae and the passerines. The order here follows Jarvis et al.
The main division in Telluraves is between the group containing the hawks, owl, and woodpeckers (Afroaves) and the group containing the falcons, parrots, and passerines (Australaves) (Ericson et al., 2006a; Hackett et al., 2008; Ericson, 2012; Jarvis et al., 2014). Australaves is by far the larger group, containing about 64% of all bird species. In contrast, Afroaves only has about 11½% of the species. Still, Afroaves is much larger than any other high-level group than branches off sooner.
I'm not fond of the names Afroaves and Australaves, mainly because the names suggest some knowledge of their origin that we don't have. As you will note in some of comments in the next few pages, some of the groups where the crown group appears to originate in Africa or Australia today (Ericson, 2012) have fossil records that are quite different. The earliest fossils throughout the clade come from the Northern Hemisphere. More precisely, they come from Eupore and North America, which were connected in Paleocene and early Eocene.
Afroaves Ericson, 2012
In the TiF list, Afroaves is divided into three groups: Accipitrimorphae, Strigiformes, and Picimorphae. There is some uncertainty about whether Strigiformes are closer to Accipitrimorphae or to Picimorphae. Jarvis et al. (2014) prefer the latter, and that is followed here.
ACCIPITRIMORPHAE Vieillot, 1816
CATHARTIFORMES Coues, 1884
Although New World vultures are rather similar to Old World Vultures (part of Accipitridae), they are not closely related. This was already recognized by Huxley in 1876 based on morphological evidence. Nonetheless, they were traditionally placed in Falconiformes, as where the Accipitridae and Falconidae. There are also some superficial similarities to storks, and when Sibley and Ahlquist found support for this using DNA hybridization techniques, they were removed from Falconiformes and placed next to the storks. More recent analysis have shown this is incorrect, and have also broken up Falconiformes. As mentioned above, more recent research (e.g., Hackett et al., 2008; Han et al., 2011; McCormack et al., 2013; Yuri et al., 2013; Jarvis et al., 2014) puts the New World vultures close to the Accipitriformes.
Cathartidae: New World Vultures de Lafresnaye, 1839
5 genera, 7 species HBW-2
- Turkey Vulture, Cathartes aura
- Lesser Yellow-headed Vulture, Cathartes burrovianus
- Greater Yellow-headed Vulture, Cathartes melambrotus
- Black Vulture, Coragyps atratus
- King Vulture, Sarcoramphus papa
- California Condor, Gymnogyps californianus
- Andean Condor, Vultur gryphus
ACCIPITRIFORMES Vieillot, 1816
Those interested in ancient raptors should read Darren Naish's post on Titan-Hawks.
The Secretarybird is the first branch of Accipitriformes, followed by the Osprey. After that, come the Accipitridae (hawks, kites, and eagles).
Sagittariidae: Secretarybird Finsch & Hartlaub, 1870 (1825)
1 genus, 1 species HBW-2
- Secretarybird, Sagittarius serpentarius
Pandionidae: Osprey Bonaparte, 1854
1 genus, 1 species HBW-2
It is not clear whether the Osprey is a single species. The IOC recognizes two Osprey species, Eastern (Indo-Australasian cristatus) and Western (everything else). Some have suggested that the Caribbean population (ridgwayi) should also be separated.
The TiF list does not currently accept either of these splits. Monti et al. (2015) sampled over 200 individuals world-wide. They found 4 clades: New world (carolinensis, including ridgwayi), Old World (haliaetus), Indo-Australian (cristatus), and North-East Asia (name uncertain). The North-East Asian clade included samples from Japan and the Pacific coast of Siberia, as well as a few from Indonesia/New Guinea and one from East Asia. Also, a cristatus haplotype was found in India. At first glance, it seems friedmanni may apply to the NE Asian clade, but Wolfe's description of friedmanni (Auk, Oct., 1946) states that Ospreys from Japan are not friedmani, making me wonder which clade the type of friedmanni actually belongs to.
Monti et al. suggest that the New World clade is basal, as suggested by the cyt-b analysis. However, combining ND2 with cyt-b yielded a different topology, with the Old World clade basal. In any event, the separation of all 4 clades appears to have taken place over a short time interval. The cyt-b analysis suggested the New World clade is a million or so years old. A few of the samples showed a mismatch between geography and clade.
At present, it remains unclear whether or not the Osprey should be considered a considered a single species, or 3-4.
- Osprey, Pandion haliaetus
Accipitridae: Hawks, Kites, Eagles Vigors, 1824
73 genera, 252 species HBW-2
At the genus level, the order presented here attempts to synthesize the papers by Amaral et al. (2006, 2009), Barrowclough et al. (2014), Breman et al. (2013), Griffiths et al. (2007), Haring et al. (2007b), Helbig et al. (2005), Kocum (2006), Lerner and Mindell (2005), Lerner et al. (2008), Nagy and Tökölyi (2014), Oatley et al. (2015), Ong et al. (2011), and Wink and Sauer-Gürth (2004). Some of the genera were restructured based on these and other papers.
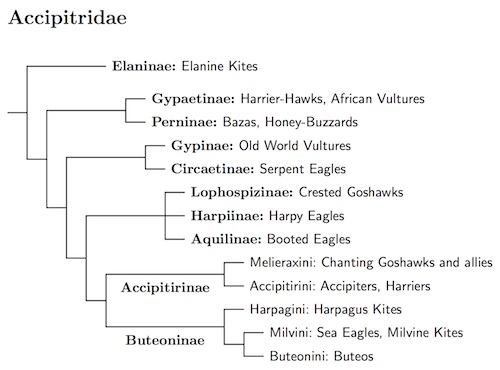
The Accipitridae are a complicated family. You can see from the diagram that I treat them as consisting of ten subfamilies with large families Accipitrinae and Buteoninae at the end. The latter two subfamilies are further divided into tribes. The timing estimates of Nagy and Tökölyi (2014) have influenced what gets counted as a subfamily and what counts as a tribe. After reading Jarvis et al. (2014), I suspect Nagy and Tökölyi's divergence dates are a little short. Nagy and Tökölyi did not include the crested goshawk subfamily, Lophospizinae, but indications are that it is near Aquilinae (Oatley et al., 2015).
The subfamily Accipitrinae is slightly smaller than Buteoninae, so I put it ahead of Buteoninae. The crested goshawks seem to be basal to both of them. There's now some evidence they belong in the Harpiinae plus Aquilinae group, which is sister to the Accipitrinae + Buteoninae group. The grouping of these five families is supported by morphology as well as genetics. Mayr (2014b) found that four of them (with the fifth not tested) share a derived type of radial carpal bone. The closest relatives of this big clade are Circaetinae and Gypinae. All together are sister to Gypaetinae and Perninae, and the whole shebang is sister to the basal group, Elaninae. This is all summed up in the diagram.
 |
| Click for genus-level tree of Accipitriformes |
|---|
Elaninae: Elanine Kites Blyth, 1851
One thing that genetic results have made clear is that the kites are not a natural group. Rather, they are scattered in three subfamilie. The first group is the Elanine kites. Kites also occur in three clades in Perninae and four clades in Buteoninae.
- Pearl Kite, Gampsonyx swainsonii
Click for Accipitridae
species tree - Scissor-tailed Kite, Chelictinia riocourii
- Black-winged Kite, Elanus caeruleus
- Black-shouldered Kite, Elanus axillaris
- Letter-winged Kite, Elanus scriptus
- White-tailed Kite, Elanus leucurus
Gypaetinae: African Vultures, Harrier-Hawks Bonaparte, 1831
Now that it is clear that Henicopernis belongs within Perninae (Barrowclough et al., 2014), this small primarily African clade is cleanly separated from Perninae. The separation from Perninae seems to be quite ancient (Nagy and Tökölyi, 2014), so I recognize both groups as subfamilies.
- African Harrier-Hawk, Polyboroides typus
- Madagascan Harrier-Hawk, Polyboroides radiatus
- Palm-nut Vulture, Gypohierax angolensis
- Egyptian Vulture, Neophron percnopterus
- Bearded Vulture / Lammergeier, Gypaetus barbatus
Perninae: Bazas, Honey-Buzzards Blyth, 1851
The Honey-Buzzards (Pernis) have been sorted out by Gamauf and Haring (2004), including the recently-split Phillipine Honey-Buzzard. Barrowclough et al. (2014) provided additional information concerning the Australo-Papuan species.
Three groups of kites are part of Perninae. The Neotropical Chondrohierax and Leptodon kites, the Swallow-tailed Kite (Elanoides, sister to the Pernis honey-buzzards), and the Square-tailed Kite (Lophoictinia, sister to the Henicopernis honey-buzzards). Note that the honey-buzzards are not a natural group either.
- Madagascan Serpent-Eagle, Eutriorchis astur
- Hook-billed Kite, Chondrohierax uncinatus
- Gray-headed Kite, Leptodon cayanensis
- White-collared Kite, Leptodon forbesi
- Swallow-tailed Kite, Elanoides forficatus
- European Honey-Buzzard, Pernis apivorus
- Philippine Honey-Buzzard, Pernis steerei
- Oriental Honey-Buzzard / Crested Honey-Buzzard, Pernis ptilorhynchus
- Barred Honey-Buzzard, Pernis celebensis
- Black-breasted Buzzard, Hamirostra melanosternon
- Square-tailed Kite, Lophoictinia isura
- Long-tailed Honey-Buzzard, Henicopernis longicauda
- Black Honey-Buzzard, Henicopernis infuscatus
- Black Baza, Aviceda leuphotes
- African Cuckoo-Hawk, Aviceda cuculoides
- Madagascan Cuckoo-Hawk, Aviceda madagascariensis
- Jerdon's Baza, Aviceda jerdoni
- Pacific Baza, Aviceda subcristata
Gypinae: Old World Vultures Blyth, 1851
The name Gypinae (Blyth, 1851) has priority over Aegypiinae (W.P. Sclater, 1924). The Old World Vultures are gathered here and fall into two clades. The first four species (and genera!) are in one clade, with Necrosyrtes and Gyps in the other. The arrangement of species within Gyps has taken both Arshad et al. (2009) and Nagy and Tökölyi (2014) into account. I follow Lerner et al. (2005) and Nagy and Tökölyi (2014) for the genera. Arshad et al. is almost the same, while Griffiths et al. (2009) is broadly consistent but less well-resolved.
- Red-headed Vulture, Sarcogyps calvus
- White-headed Vulture, Trigonoceps occipitalis
- Cinereous Vulture, Aegypius monachus
- Lappet-faced Vulture, Torgos tracheliotos
- Hooded Vulture, Necrosyrtes monachus
- White-backed Vulture, Gyps africanus
- White-rumped Vulture, Gyps bengalensis
- Himalayan Vulture, Gyps himalayensis
- Griffon Vulture, Gyps fulvus
- Rueppell's Vulture, Gyps rueppelli
- Cape Vulture, Gyps coprotheres
- Slender-billed Vulture, Gyps tenuirostris
- Indian Vulture, Gyps indicus
Circaetinae: Serpent Eagles Blyth, 1851
In the Serpent Eagles, Dryotriorchis has been merged into Circaetus. Exactly how many species are in Spilornis remains an issue. There are six here, but Ferguson-Lees and Christie (2001) list 13! I've not accepted the IOC split of minimus because I don't see any evidence to split this and not the others.
- Andaman Serpent-Eagle, Spilornis elgini
- Philippine Serpent-Eagle, Spilornis holospilus
- Crested Serpent-Eagle, Spilornis cheela
- Great Nicobar Serpent-Eagle, Spilornis klossi
- Mountain Serpent-Eagle, Spilornis kinabaluensis
- Sulawesi Serpent-Eagle, Spilornis rufipectus
- Philippine Eagle, Pithecophaga jefferyi
- Bateleur, Terathopius ecaudatus
- Western Banded Snake-Eagle, Circaetus cinerascens
- Southern Banded Snake-Eagle, Circaetus fasciolatus
- Congo Serpent-Eagle, Circaetus spectabilis
- Beaudouin's Snake-Eagle, Circaetus beaudouini
- Black-chested Snake-Eagle, Circaetus pectoralis
- Brown Snake-Eagle, Circaetus cinereus
- Short-toed Snake-Eagle, Circaetus gallicus
Lophospizinae: Crested Goshawks Informal
What evidence there is suggests that the Crested Goshawk is not part of Accipitrinae. Barcoding suggested it is basal to the Accipitrinae/Buteoninae clade (Ong et al., 2011; Breman et al., 2013). Raty, in a BirdForum post got similar results when adding a partial cytochrome-b gene. More recently, Oatley et al.'s (2015) 4-gene analysis found the Crested Goshawk near the Aquilinae (Harpiinae was not in the analysis). I'm not sure I'm entirely convinced, but I've double-down on this and put it in a trichotomy with Harpiinae and Aquilinae. Besides, there's something that appeals to me in having the crested goshawks near the harpy eagles.
DNA from the Sulawesi (Crested) Goshawk has not been tested, but it is thought to be a very close relative of the Crested Goshawk, with the two forming a superspecies. The name Lophospiza (Kaup 1844, type trivirgata) is available for these two species.
- Crested Goshawk, Lophospiza trivirgata
- Sulawesi Goshawk, Lophospiza griseiceps
Harpiinae: Harpy Eagles Lesson, 1828
The split between the harpy eagles and booted eagles seems to have occurred about 20 million years ago (Nagy and Tökölyi, 2014), so I treat them as separate subfamilies.
- Bat Hawk, Macheiramphus alcinus
- Papuan Eagle, Harpyopsis novaeguineae
- Harpy Eagle, Harpia harpyja
- Crested Eagle, Morphnus guianensis
Aquilinae: Booted Eagles Vigors, 1825
The key papers for the Booted Eagles (Aquilinae) are Bunce et al. (2005),
Helbig et al. (2005), Lerner and Mindell (2005), and Haring et al.
(2007b).
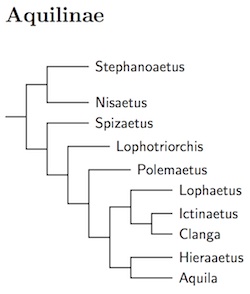 The Spizaetus Hawk-Eagles belong in two different clades within
Aquilinae. Thus Spizaetus is divided into Nisaetus and
Spizaetus. The Black-and-chestnut Eagle (Oroaetus) must
be merged into the remaining Spizaetus. The Rufous-bellied Hawk-Eagle
(Lophotriorchis) is separated from Hieraaetus. while
the spotted eagles are separated from Aquila as Clanga.
Hieraaetus loses a couple of species to Aquila, which also
gains Cassin's Hawk-Eagle from Spizaetus.
The Spizaetus Hawk-Eagles belong in two different clades within
Aquilinae. Thus Spizaetus is divided into Nisaetus and
Spizaetus. The Black-and-chestnut Eagle (Oroaetus) must
be merged into the remaining Spizaetus. The Rufous-bellied Hawk-Eagle
(Lophotriorchis) is separated from Hieraaetus. while
the spotted eagles are separated from Aquila as Clanga.
Hieraaetus loses a couple of species to Aquila, which also
gains Cassin's Hawk-Eagle from Spizaetus.
I've included the extinct Haast's Eagle of New Zealand on the species tree since Bunce et al. (2005) analyzed DNA from it. However, it isn't included on the main list because it likely became extinct prior to 1500, which is the cutoff for inclusion.
- Crowned Eagle, Stephanoaetus coronatus
- Sulawesi Hawk-Eagle, Nisaetus lanceolatus
- Pinsker's Hawk-Eagle, Nisaetus pinskeri
- Changeable Hawk-Eagle, Nisaetus cirrhatus
- Philippine Hawk-Eagle, Nisaetus philippensis
- Wallace's Hawk-Eagle, Nisaetus nanus
- Mountain Hawk-Eagle, Nisaetus nipalensis
- Legge's Hawk-Eagle, Nisaetus kelaarti
- Blyth's Hawk-Eagle, Nisaetus alboniger
- Javan Hawk-Eagle, Nisaetus bartelsi
- Black Hawk-Eagle, Spizaetus tyrannus
- Black-and-white Hawk-Eagle, Spizaetus melanoleucus
- Ornate Hawk-Eagle, Spizaetus ornatus
- Black-and-chestnut Eagle, Spizaetus isidori
- Rufous-bellied Hawk-Eagle, Lophotriorchis kienerii
- Martial Eagle, Polemaetus bellicosus
- Long-crested Eagle, Lophaetus occipitalis
- Black Eagle, Ictinaetus malaiensis
- Lesser Spotted-Eagle, Clanga pomarina
- Greater Spotted-Eagle, Clanga clanga
- Indian Spotted-Eagle, Clanga hastata
- Wahlberg's Eagle, Hieraaetus wahlbergi
- Ayres's Hawk-Eagle, Hieraaetus ayresii
- Pygmy Eagle, Hieraaetus weiskei
- Booted Eagle, Hieraaetus pennatus
- Little Eagle, Hieraaetus morphnoides
- Steppe Eagle, Aquila nipalensis
- Tawny Eagle, Aquila rapax
- Spanish Imperial-Eagle, Aquila adalberti
- Eastern Imperial-Eagle, Aquila heliaca
- Golden Eagle, Aquila chrysaetos
- Cassin's Hawk-Eagle, Aquila africana
- Gurney's Eagle, Aquila gurneyi
- Wedge-tailed Eagle, Aquila audax
- Verreaux's Eagle, Aquila verreauxii
- Bonelli's Eagle, Aquila fasciata
- African Hawk-Eagle, Aquila spilogaster
Accipitrinae: Accipiters, Harriers Vigors, 1824
There is a deep division of the Accipitrinae into two groups which I rank as tribes: Melieraxini and Accipitrini.
Melieraxini: Chanting Goshawks and allies Informal
The Tiny Hawk (and presumably Semicollared Hawk) do not seem closely related to Accipiter (Kocum, 2006; Olson, 2006) and have been removed from Accipiter and placed in the genus Hieraspiza (Kaup 1844, type superciliosa). It has not been clear what they are related to.
Kocum (2006) includes several species from this group in her dissertation. Her analysis places them in a weakly supported unresolved basal polytomy with Accipitrini, Harpagini, and the remaining Buteoninae. Griffiths et al. (2007) grouped together the Lizard Buzzard, Gabar Goshawk, Long-tailed Hawk, and the chanting goshawks. The group was basal to Accipitrini and Buteoninae, but did not include the Tiny Hawk. The analysis by Nagy and Tökölyi (2014) bundles them together as shown here, but their evidence for this seems weak.
The name Melieraxinae has appeared in the literature since at least Lerner and Mindell (2005), but does not seem to be formally established.
- Lizard Buzzard, Kaupifalco monogrammicus
- Gabar Goshawk, Micronisus gabar
- Long-tailed Hawk, Urotriorchis macrourus
- Tiny Hawk, Hieraspiza superciliosa
- Semicollared Hawk, Hieraspiza collaris
- Dark Chanting-Goshawk, Melierax metabates
- Eastern Chanting-Goshawk, Melierax poliopterus
- Pale Chanting-Goshawk, Melierax canorus
The papers by Wink and Sauer-Gürth (2004), Breman et al. (2013), Barrowclough et al. (2014), Nagy and Tökölyi (2014) and Kocum's dissertation (2006) allow us to make a rough draft of a species-level tree for the accipitters. Doubtless it will need further adjustment.
The result is that the genus Accipiter has been divided into 6 pieces. Two of those, Hieraspiza and Lophospiza have been moved outside the Accipitrini. Kocum (2006), Griffiths et al. (2007), Lerner et al. (2008), and Nagy and Tökölyi (2014) found that the Circus harriers are embedded within the traditional Accipiter. The options are to lump Circus into Accipiter or divide Accipiter into at least three parts. I've chosen to take the latter course, dividing Accipter into four parts.
The Cooper's Hawk, Northern Goshawk, and closely related taxa are the closest relatives of the harriers. They go in genus Astur (Lacépède 1799, type gentilis). These are sister to a clade that includes the Eurasian Sparrowhawk and the Sharp-shinned Hawk complex. It retains the name Accipiter (Brisson 1760, type nisus). I have included the Gray-bellied Hawk with this group based on Kocum (2006), although the support for this is not strong.
The remaining accipiters and Erythrotriorchis form a clade sister to the combined Accipiter-Astur-Circus clade. There is a deep division (ca. 15 million years) between a small group of African accipiters and the remaining accipiters, so they are divided into two genera: Aerospiza (Roberts 1922, type tachiro) and Tachyspiza. There are two oldest available names for the Tachyspiza clade: Tachyspiza (type soloensis) and Leucospiza (type novaehollandiae), both from the same 1844 monograph by Kaup. Neither seems to have obtained any priority over the other, and I'm somewhat arbitrarily using Tachyspiza here. However, someone will need to formally pick one in the literature and give it priority, hopefully Tachyspiza. The name Leucospiza really only fits the white morph of novaehollandiae. At present, it is unclear where exactly Erythrotriorchis fits in. It may be sister to Aerospiza, or to Tachyspiza or even embedded within one of them (Barrowclough et al, 2014 is not conclusive).
Within Tachyspiza, the position of the Spot-tailed Sparrowhawk, Tachyspiza trinotata, is quite uncertain. I also have doubts about where the henicogramma-novaehollandiae clade goes. Nagy and Tökölyi (2014), apparently using the RAG-1 gene from subspecies rufoschistaceus (hiogaster, not novaehollandiae) put it near imitator, but Breman et al. (2013), using the barcoding region, put another member of the clade, fasciata, nearer virgata. There is also some uncertainty about exactly where soloensis goes. I've followed Kocum's (2006) multigene analysis, but Nagy and Tökölyi (2014) have it in a rather different position.
This list includes three relatives of the Sharp-shinned Hawk that SACC has not split (although they are split in the Sibley-Monore list). They could be allopatric forms best treated as one species. However, although the combined analysis of Kocum (2006) placed ventralis and erythronemius in a clade striatus, other arrangements were seen for some of the other genes (chionogaster was not included). Breman et al. (2013) also found a separation between striatus and erythronemius. This hints, but does not prove, that a species-level treatment is appropriate, as has been followed by various authors.
The arrangement of the harriers follows Oatley et al. (2015). I have split the Hen and Northern Harriers. Oatley et al. found that the Northern Harrier was closer to the Cinereous Harrier than to the Hen Harrier. Oatley et al. also found that the last 5 harriers are very closely related, possibily conspecific. However, I distrust their estimated divergence dates. They are markedly shorter than comparable dates in Nagy and Tökölyi (2014), which already seem a bit short compared to Jarvis et al. (2015).
Accipitrini: Accipiters, Harriers
- Chestnut-shouldered Goshawk, Erythrotriorchis buergersi
- Red Goshawk, Erythrotriorchis radiatus
- Chestnut-flanked Sparrowhawk, Aerospiza castanilius
- Red-chested Goshawk, Aerospiza toussenelii
- African Goshawk, Aerospiza tachiro
- Red-thighed Sparrowhawk, Tachyspiza erythropus
- Little Sparrowhawk, Tachyspiza minulla
- Spot-tailed Sparrowhawk, Tachyspiza trinotata
- Japanese Sparrowhawk, Tachyspiza gularis
- Besra, Tachyspiza virgata
- Dwarf Sparrowhawk, Tachyspiza nana
- Vinous-breasted Sparrowhawk, Tachyspiza rhodogaster
- Rufous-necked Sparrowhawk, Tachyspiza erythrauchen
- Collared Sparrowhawk, Tachyspiza cirrocephala
- New Britain Sparrowhawk, Tachyspiza brachyura
- Moluccan Goshawk, Tachyspiza henicogramma
- Brown Goshawk, Tachyspiza fasciata
- Variable Goshawk, Tachyspiza hiogaster
- Gray Goshawk, Tachyspiza novaehollandiae
- Shikra, Tachyspiza badia
- Levant Sparrowhawk, Tachyspiza brevipes
- Nicobar Sparrowhawk, Tachyspiza butleri
- Chinese Sparrowhawk, Tachyspiza soloensis
- Black-mantled Goshawk, Tachyspiza melanochlamys
- Pied Goshawk, Tachyspiza albogularis
- White-bellied Goshawk, Tachyspiza haplochroa
- Fiji Goshawk, Tachyspiza rufitorques
- Frances's Sparrowhawk, Tachyspiza francesiae
- Slaty-mantled Goshawk, Tachyspiza luteoschistacea
- Imitator Goshawk, Tachyspiza imitator
- Gray-headed Goshawk, Tachyspiza poliocephala
- New Britain Goshawk, Tachyspiza princeps
- Gray-bellied Hawk, Accipiter poliogaster
- Ovambo Sparrowhawk, Accipiter ovampensis
- Madagascan Sparrowhawk, Accipiter madagascariensis
- Eurasian Sparrowhawk, Accipiter nisus
- Rufous-breasted Sparrowhawk, Accipiter rufiventris
- Sharp-shinned Hawk, Accipiter striatus
- White-breasted Hawk, Accipiter chionogaster
- Plain-breasted Hawk, Accipiter ventralis
- Rufous-thighed Hawk, Accipiter erythronemius
- Doria's Goshawk, Megatriorchis doriae
- Bicolored Hawk, Astur bicolor
- Cooper's Hawk, Astur cooperii
- Gundlach's Hawk, Astur gundlachi
- Northern Goshawk, Astur gentilis
- Meyer's Goshawk, Astur meyerianus
- Black Sparrowhawk, Astur melanoleucus
- Henst's Goshawk, Astur henstii
- Spotted Harrier, Circus assimilis
- Pallid Harrier, Circus macrourus
- Black Harrier, Circus maurus
- Hen Harrier, Circus cyaneus
- Northern Harrier, Circus hudsonius
- Cinereous Harrier, Circus cinereus
- Long-winged Harrier, Circus buffoni
- Montagu's Harrier, Circus pygargus
- Pied Harrier, Circus melanoleucos
- Western Marsh-Harrier, Circus aeruginosus
- African Marsh-Harrier, Circus ranivorus
- Swamp Harrier, Circus approximans
- Papuan Harrier, Circus spilothorax
- Reunion Harrier, Circus maillardi
- Eastern Marsh-Harrier, Circus spilonotus
- Malagasy Harrier, Circus macrosceles
Buteoninae: Buteos Vigors 1824
The Buteoninae are the largest subfamily of the Accipitridae. According to Nagy and Tökölyi (2014), the clade is roughly 20 million years old. There are three deep divisions (15-18 million years ago) in the Buteoninae which I have recognized as tribes: Harpagini, Milvini, and Buteonini.
Many of the changes between versions 2.00 and 2.01 of this page were due to the publication of Lerner et al. (2008). Their results have subsequently been refined by Amaral et al. (2009) and Nagy and Tökölyi (2014). I have additionally consulted Amaral et al. (2006), Barrowclough et al.\ (2014), Griffiths et al.\ (2007), Kocum (2006), Kruckenhauser et al.\ (2004), Ong et al.\ (2011), and Riesing et al. (2003). Although these sources are not in 100% agreement, and there are still a few lacuna, I think the Buteoninae are now in pretty good shape even at the species level.
There might still be a little modification of species boundaries to go, particularly in Pseudastur and near Buteo buteo. I've modified the generic limits quite a bit in the Buteoninae, but less drastically than suggested by Riesing et al. (2003) and by Lerner et al. (2008). Most of these changes were also adopted by Amaral et al. (2009), which prompted some further changes in version 2.15. This has been further refined by considering Nagy and Tökölyi (2014) in version 2.61.
Harpagini Bonaparte, 1854
Harpagini consists of the two Harpagus kites. Nagy and Tökölyi (2014) put Harpagini in the basal position. Griffiths et al. (2007) also included them in their analysis. They have Harpagus basal to Accipitrinae + Buteoninae. However, resolution was poor, and other basal raptors from Griffiths et al. have moved to Accipitridae.
- Double-toothed Kite, Harpagus bidentatus
- Rufous-thighed Kite, Harpagus diodon
Milvini: Sea Eagles and Milvine Kites Vigors 1824
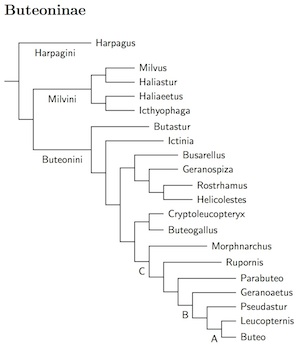 Milvini consists of the milvine kites and sea and fish eagles. The
kites are the second of the four kite clades in Buteoninae.
Milvini consists of the milvine kites and sea and fish eagles. The
kites are the second of the four kite clades in Buteoninae.
The two-species version of Icthyophaga turns out to be nested within Haliaeetus. It could either be submerged into Haliaeetus, or expanded to include four Haliaeetus species. I've taken the second option. This puts all of the northern fish-eagles and sea-eagles in Haliaeetus, while the tropical fish/sea-eagles are in Icthyophaga. The two groups are not only geographically separated, but are visually distinct—compare the relatively longer, narrower wings of Haliaeetus to the shorter, broader wings of Icthyophaga. There's a spelling issue here: Icthyophaga or Ichthyophaga? Both Dickinson (2003) and Peterson (zoonomen.net) give the former, which is the original spelling. Peterson also argues that Icthyophaga is correct. Nonetheless, Ichthyophaga is also in wide usage (e.g., HBW-2).
- Brahminy Kite, Haliastur indus
- Whistling Kite, Haliastur sphenurus
- Red Kite, Milvus milvus
- Black Kite, Milvus migrans
- Yellow-billed Kite, Milvus aegyptius
- Pallas's Fish-Eagle, Haliaeetus leucoryphus
- Steller's Sea-Eagle, Haliaeetus pelagicus
- White-tailed Eagle, Haliaeetus albicilla
- Bald Eagle, Haliaeetus leucocephalus
- White-bellied Sea-Eagle, Icthyophaga leucogaster
- Sanford's Sea-Eagle, Icthyophaga sanfordi
- African Fish-Eagle, Icthyophaga vocifer
- Madagascan Fish-Eagle, Icthyophaga vociferoides
- Lesser Fish-Eagle, Icthyophaga humilis
- Gray-headed Fish-Eagle, Icthyophaga ichthyaetus
Buteonini: Buteos Vigors 1824
This brings us to Buteonini. The first branch consists of Butastur buzzards and the second branch is the Ictinia kites (third of the four Buteoninae kite clades). The next clade contains Busarellus, Geranospiza, and the final kite clade, Rostrhamus and presumably Helicolestes. Except possibly for the last two, they are all long-separated and deserving of genus status.
There have been big changes for Buteogallus. The traditional Buteogallus hawks turned out to be closely related to Harpyhaliaetus and three of the Leucopternis hawks. Prior to version 2.15, I was rolling them all into Buteogallus. Then, I followed the recommendations of Amaral et al. (2009), who create two new genera and also slightly adjust the taxonomy. After several rounds of debate, the SACC has settled on another solution, which I follow as of version 2.55. They recognize the new genus Cryptoleucopteryx (Amaral et al, 2009) for the Plumbeous Hawk (formerly in Leucopternis). Buteogallus now applies to the old Buteogallus plus Slate-colored Hawk and White-necked Hawk (from Leucopternis) and the two Harpyhaliaetus solitary-eagles. Amaral et al. (2009) and Nagy and Tökölyi (2014) estimate the common Buteogallus ancestor at about 4-6 million years ago, so putting them all in one genus is reasonable. In contrast, they estimate that the original Buteogallus separated from Cryptoleucopteryx somewhere between 7 and 10 million years ago.
This brings us to point C, one place where Lerner et al. suggest the rest be considered Buteo. I think this idea hides more taxonomy than it reveals and prefer a somewhat different arrangement. As it happens, Amaral et al. (2009) came to similar conclusions. Amaral et al. and Nagy and Tökölyi (2014) estimate that Morphnarchus separated from Buteo about 8-11 million years ago. Morphnarchus is another Leucopternis refugee, while the Roadside Hawk (Rupornis) is sometimes considered part of Buteo. The same is true of the White-rumped Hawk, which has joined Harris's Hawk in Parabueto. Nagy and Tökölyi (2014) and Barrowclough et al. (2014) put Rupornis in a clade with Parabueto, but I believe that Amaral et al. (2009) are used more data here and found them on sequential branches.
The clade starting at B is also flagged by Lerner et al. as a good starting point for Buteo. The former Leucopternis, White, Gray-backed, and Mantled Hawks are placed in Pseudastur while the some of the remaining Leucopternis are put in Geranoaetus. This includes the White-tailed, and Variable (sometimes split as Red-backed and Puna) Hawks, formerly in Buteo. It should be noted that the species boundaries in Pseudastur seriously need adjustment, but it seems that further study will be needed to clarify the situation.
The clade at C is also a plausible way to delimit Buteo. The SACC prefers to retain Leucopternis. The balance of evidence suggests Leucopternis is more basal than the Gray and Gray-lined Hawks (subgenus Asturina). Amaral et al. (2009) find Asturina sister to Buteo and Nagy and Tökölyi (2014) put the Asturina hawks a little deeper in the buteos. I now follow both AOU committees by including Asturina in Buteo.
Based on Millsap et al. (2011), the Gray Hawk, formerly Asturina nitida, has been split into Gray Hawk, Asturina plagiata and Gray-lined Hawk, Asturina nitida.
As defined below, Buteo consists primarily of species breeding in the Old World and Nearctic, with a few Neotropical species. The other genera that might be included in Buteo — Leucopternis (point A), Pseudastur, Geranoaetus (point B), Parabuteo, Rupornis, and Morphnarchus (point C) — are primarily South and Middle American breeders.
Amaral et al. (2009) and Nagy and Tökölyi (2014) help clear up the situation with the remaining Buteo species, although some issues remain. The New World species split out nicely, up to the lagopus/regalis pair. Then we get into the Old World buteos, whose taxonomy remains somewhat murky. Two species were not considered in their analysis, and the true species boundaries among the Old World buteos remain somewhat uncertain. I expect that the treatment here will subject to a bit of revision as more is known.
Riesing et al. (2003, Fig.5) found evidence that Archer's Buzzard, Buteo archeri is not part of the Augur Buzzard, Buteo augur, but is sister to a clade consisting of the Red-necked, Augur and Jackal Buzzards, and possibly the Madagascan Buzzard, Buteo brachypterus. Other evidence in Riesing et al. (Fig.4), and in Nagy and Tökölyi (2014) puts brachypterus in a slightly more basal position, out of this clade.
Another apparently misplaced buteo was the Forest Buzzard, Buteo trizonatus. It was considered a subspecies of the Mountain Buzzard, Buteo oreophilus, but genetic data (esp. Kruckenhauser et al., 2004) suggests it is closer to the Common Buzzard, Buteo buteo. Although the genetic distance is small, its distant separation from the Common Buzzard's breeding range and distinct plumage suggest it should be considered a distinct species.
Although the breeding buzzards of the Cape Verde and Socotra Islands have been considered races of the Common Buzzard, genetic data (Clouet and Wink, 2000) suggests they are more closely related to the Long-legged Buzzard, Buteo rufinus. Once again, the genetic distances are small, but this suggests treating them as distinct species: Cape Verde Buzzard, Buteo bannermani, and Socotra Buzzard, Buteo socotraensis. Oddly, although it had been studied for over a century, the Socotra Buzzard had not been formally named until 2010 when Porter and Kirwan dubbed it Buteo socotraensis. This replaces the informal name ‘B. socotrae’ that some have used.
- Grasshopper Buzzard, Butastur rufipennis
- White-eyed Buzzard, Butastur teesa
- Rufous-winged Buzzard, Butastur liventer
- Gray-faced Buzzard, Butastur indicus
- Mississippi Kite, Ictinia mississippiensis
- Plumbeous Kite, Ictinia plumbea
- Black-collared Hawk, Busarellus nigricollis
- Crane Hawk, Geranospiza caerulescens
- Snail Kite, Rostrhamus sociabilis
- Slender-billed Kite, Helicolestes hamatus
- Plumbeous Hawk, Cryptoleucopteryx plumbea
- Slate-colored Hawk, Buteogallus schistaceus
- Rufous Crab Hawk, Buteogallus aequinoctialis
- Common Black Hawk, Buteogallus anthracinus
- Cuban Black Hawk, Buteogallus gundlachii
- Savanna Hawk, Buteogallus meridionalis
- White-necked Hawk, Buteogallus lacernulatus
- Great Black Hawk, Buteogallus urubitinga
- Solitary Eagle, Buteogallus solitarius
- Chaco Eagle, Buteogallus coronatus
- Barred Hawk, Morphnarchus princeps
- Roadside Hawk, Rupornis magnirostris
- Harris's Hawk, Parabuteo unicinctus
- White-rumped Hawk, Parabuteo leucorrhous
- White-tailed Hawk, Geranoaetus albicaudatus
- Variable Hawk, Geranoaetus polyosoma
- Black-chested Buzzard-Eagle, Geranoaetus melanoleucus
- Gray-backed Hawk, Pseudastur occidentalis
- White Hawk, Pseudastur albicollis
- Mantled Hawk, Pseudastur polionotus
- Semiplumbeous Hawk, Leucopternis semiplumbeus
- Black-faced Hawk, Leucopternis melanops
- White-browed Hawk, Leucopternis kuhli
- Gray Hawk, Buteo plagiatus
- Gray-lined Hawk, Buteo nitidus
- Red-shouldered Hawk, Buteo lineatus
- Ridgway's Hawk, Buteo ridgwayi
- Broad-winged Hawk, Buteo platypterus
- Hawaiian Hawk, Buteo solitarius
- White-throated Hawk, Buteo albigula
- Short-tailed Hawk, Buteo brachyurus
- Galapagos Hawk, Buteo galapagoensis
- Swainson's Hawk, Buteo swainsoni
- Zone-tailed Hawk, Buteo albonotatus
- Red-tailed Hawk, Buteo jamaicensis
- Rufous-tailed Hawk, Buteo ventralis
- Rough-legged Hawk / Rough-legged Buzzard, Buteo lagopus
- Ferruginous Hawk, Buteo regalis
- Madagascan Buzzard, Buteo brachypterus
- Archer's Buzzard, Buteo archeri
- Red-necked Buzzard, Buteo auguralis
- Augur Buzzard, Buteo augur
- Jackal Buzzard, Buteo rufofuscus
- Eastern Buzzard, Buteo japonicus
- Upland Buzzard, Buteo hemilasius
- Himalayan Buzzard, Buteo burmanicus
- Mountain Buzzard, Buteo oreophilus
- Common Buzzard, Buteo buteo
- Forest Buzzard, Buteo trizonatus
- Long-legged Buzzard, Buteo rufinus
- Cape Verde Buzzard, Buteo bannermani
- Socotra Buzzard, Buteo socotraensis